CIENCIAS AGROPECUARIAS-Artículo Científico
DETERMINACIÓN DE LA HABILIDAD COMPETITIVA ENTRE Daucus carota L. Y Senecio vulgaris L.
DETERMINATION OF COMPETITIVE ABILITY BETWEEN Daucus carota L. AND Senecio vulgaris L.
Mónica Dotor Robayo1, Marco Cabezas Gutiérrez2
1Ingeniera Agrónoma, Maestría Fisiología Vegetal, Grupo Manejo Biológico de Cultivos, Facultad de Ciencias Agropecuarias. Universidad Pedagógica y Tecnológica de Colombia. Avenida Central del Norte, Tunja, Boyacá (Colombia), e-mail: monica.dotor@uptc.edu.co
2Ingeniero Agrónomo, M.Sc., cPh.D. Director Técnico Yara Colombia, Docente Maestría Fisiología Vegetal. Universidad Pedagógica y Tecnológica de Colombia. Avenida Central del Norte, Tunja, Boyacá (Colombia), e-mail:mcabezasg@unal.edu.co
Rev. U.D.C.A Act. & Div. Cient. 18(1): 81-89, Enero-Junio, 2015
RESUMEN
Desde la perspectiva agronómica, conocer el nivel de competencia entre una especie cultivada y las plantas invasoras permite prever las pérdidas en rendimiento y establecer la importancia del manejo adecuado de las especies invasoras, durante el proceso productivo. Para contribuir a la comprensión de este proceso, se realizó un ensayo que permitiera advertir la capacidad competitiva entre las especies Senecio vulgaris L. y Daucus carota L. El experimento, se realizó bajo un diseño completamente al azar, con cuatro tratamientos y cinco repeticiones, para identificar el efecto individual y combinado de los factores luz y agua, sobre el desarrollo y la producción de estas especies. Se observó que cuando D. carota crece en condiciones de competencia total y sin competencia muestra una relación alométrica raíz:tallo constante (4:1), pero en condiciones limitantes de agua y luz, se modifica esta relación, al parecer buscando compensar la toma del recurso limitado. Las pérdidas de peso en la especie cultivada fueron superiores en condiciones de competencia por agua, observándose un mayor porcentaje de distribución de biomasa hacia la raíz (74%), pero una menor producción y almacenamiento total, por lo que se podría suponer que este factor es crítico para la hortaliza. Se encontraron pérdidas del 43% en peso, en comparación con el tratamiento sin competencia. En el caso de S. vulgaris fue significativa la variación en la producción total de biomasa de la especie, por lo que es probable que la especie, a diferencia de la cultivada, desarrolle un patrón de ajuste en el crecimiento por competencia.
Palabras clave: Malezas, competencia inter-especifica, relaciones alométricas.
SUMMARY
From the agricultural perspective, knowing the level of competition between a crop and invasive plants can predict yield losses and establish the importance of proper management of weeds during the production process. To contribute to the understanding of this process, a test to know the competitive ability between species such as Senecio vulgaris L. and Daucus carota L. was carried out. A completely randomized experimental design with four treatments and five replicates was used in order to identify the individual and combined effect of water and light on the development and production of these species. It was observed that when growing D. carota under full competition and without competition the plant shows a root:shoot allometric relationship (4:1), but under limiting water and light conditions, this relationship changes, apparently seeking to compensate the intake of the limited resource. Losses in the crop species were higher under competition for water, showing a higher percentage of biomass allocation to the root (74%), but a lower dry biomass production and storage. It can be assumed, that this factor is critical for the vegetable, showing losses of 43% compared to the treatment without competition. In the case of S. vulgaris the variation in total biomass production was significant, so it is probable that in difference to the cultivated species, the weed develops a pattern of growth competition adjustment.
Key words: Weeds, inter-specific competition, allometric relationships.
INTRODUCCIÓN
La competencia inter-específica es un tipo de interferencia que se presenta entre diferentes especies. En el caso de la producción agrícola, se refiere a la interferencia que ocasiona una población heterogénea de plantas nocivas sobre el cultivo que, a su vez, se caracteriza por ser una población de tendencia homogénea (Begon et al. 1995; Plaza et al. 2009). Esta competencia, se genera entre especies que comparten el mismo nicho, en el cual, las plantas lidian por un factor o factores limitados y se refleja a nivel de dosel, raíz, flores, frutos, semillas (Miner et al. 2005; Ijlal et al. 2011). Normalmente, este efecto ecológico influye, de manera negativa, sobre la población menos competitiva, por lo que afecta directamente la diversidad poblacional (Begon et al. 1995; Roberts & Popai, 1970), por tanto, la habilidad para competir va a ser determinante en los procesos de sobrevivencia de las especies y en la expresión fenotípica de las plantas en oposición (Cambrón et al. 2013; Håkansson, 2003). Así, la diversidad de plantas en un ambiente y las diferentes demandas nutricionales de cada especie, contribuyen a determinar la habilidad competitiva de las mismas y a generar la tendencia natural de consolidar alguna especie como altamente competitiva y dominante en un área o un nicho específico (Acosta & Agüero, 2001; Terrazas& Germain, 1995).
Cuando una especie se encuentra limitada por un recurso busca compensar esa limitación, generando cambios fisionómicos, para los cuales, necesita modificar la asignación y la distribución de los fotoasimilados y los nutrientes (Nui et al. 2008; Vibrans, 2009). La rapidez con la que produzca los cambios y la sensibilidad a las condiciones del medio, determina su éxito como competidora y la capacidad invasiva de la especie frente a otras (Lei et al. 2013; Auld, 1996; McGinley, 2012).
Existen diferentes formas de medir los efectos de la competencia, desde la perspectiva agronómica; una opción, la constituye el análisis diferencial de producción-Yield Gap Analysis (Ittersum et al. 2013). Esta técnica consiste en realizar una evaluación del efecto que genera una condición limitante sobre la producción de un cultivo, teniendo como punto de referencia el potencial de producción máximo, de modo que sea posible reconocer el costo en rendimiento agronómico, de cada factor en evaluación (Lobell et al. 2009; Perry & Perry, 1989).
Fisiológicamente es posible determinar el efecto de una condición limitante en una especie, a través de mediciones de los diferentes componentes de rendimiento biológico o agronómico de la planta, de modo que puedan determinar los cambios en la distribución de los asimilados, según las condiciones en las que se induce el crecimiento de la misma (Camargo & Rodríguez, 2006; Qasem & Hill, 1989). Estoscambios pueden ser catalogados como de respuesta plástica fenotípica, cuando producen una variación en el patrón de asignación de los fotoasimilados en la planta y que esta respuesta es consecuencia del estímulo ambiental (Radosevich et al. 2007). Este aspecto es aplicable para el caso en que la planta se encuentre bajo una condición de recursos limitantes, siendo el principio de la teoría del equilibrio funcional (McConnaughay & Coleman, 1999; Camargo & Rodríguez, 2006). Como consecuencia, se espera que la planta intente suplir la necesidad del recurso en cuestión, modificando el proceso de traslocación, para favorecer la zona implicada en la captura del recurso, de modo que se optimice su crecimiento y este se pueda adquirir (Janeček et al. 2014; Zimdahl, 2007; Wiener, 2005).
La presente investigación, se realizó con el fin de conocer la habilidad competitiva entre Senecio vulgaris L., una de las especies invasoras de mayor impacto económico en la actualidad, en la zona hortícola de Boyacá y Daucus carota L., buscando determinar la capacidad que tienen estas especies para competir, el costo en la distribución de biomasa bajo competencia en cada una de las especies y los factores críticos que determinan el crecimiento de las especies en referencia.
MATERIALES Y MÉTODOS
Este experimento, se realizó en las instalaciones de la casa de vidrio de la Universidad Pedagógica y Tecnológica de Colombia, Sede Tunja. Como parte de la preparación preliminar del ensayo, se tomó una muestra de suelos, que fue llevada al laboratorio de suelos de la UPTC, donde los resultados mostraron un pH de 4,56, conductividad eléctrica de 0,17, acidez intercambiable 0,7, aluminio de cambio: 0,5mEq.100-1g, H 0.2, P 22,61ppm, materia orgánica 18,86%, Ca 11,02mEq.100-1g, Mg 1,43mEq.100-1g, K 3,24mEq.100-1g, Na 0,22mEq.100-1g, Fe 104mEq.100-1g, Mn 10,2ppm, Cu 0,19ppm,Zn 2,69ppm. El análisis del suelo indica que, posiblemente, corresponde a un Andisol orgánico, lo que explica el bajo nivel de pH y aluminio de cambio. A pesar de esta condición, no se realizó corrección de acidez en el suelo, para preservar las condiciones de producción de la zona.
Se utilizaron 20 materos plásticos de 3L de capacidad, llenados con suelo proveniente de la zona hortícola del municipio de Ventaquemada; los materos fueron dispuestos en un diseño experimental completamente al azar. Se asignaron cuatro tratamientos con cinco repeticiones, para buscar el efecto individual y combinado de los factores de competencia luz y agua sobre el desarrollo y la producción de la planta cultivada y la planta no cultivada, siguiendo la metodología propuesta por Doll (1989).
Descripción de tratamientos: Tratamiento 1. Competencia total. Se colocaron 20 semillas de las especies a lados opuestos del matero. La semilla de zanahoria fue hibrida Taki-Chanteney, seleccionada por ser uno de los materiales de mayor área sembrada en la región. Cuando las plantas alcanzaron un desarrollo adecuado, con hojas cotiledonales y dos hojas verdaderas, completamente desplegadas, se hizo un raleo dejando solamente un individuo por especie, que se mantuvo hasta el final del ciclo vegetativo de S. vulgaris. Con el fin de eliminar la alteración en la competencia, se retiraron las plántulas de especies diferentes a las objetivo, hasta el final del ensayo. Tratamiento 2. Competencia inter-especifica por luz. Se tomaron cinco materos y se dividió con una lámina de acetato transparente cada matero, en dos partes iguales, de modo que tuviesen independencia en el suelo; en cada parte, se agregaron cantidades similares de suelo; en cada división, se sembraron 20 semillas de una especie a un lado y 20 de la otra especie, en el lado complementario. Se procedió de manera similar que en el tratamiento 1. Tratamiento 3. Competencia inter-específica por agua y nutrientes. Se tomaron cinco materos con suelo, que fueron divididos en la parte superior con un acetato blanco, de modo que el área radicular de las especies estuviesen en un el mismo ambiente y, en la zona de crecimiento del área foliar, se mantuvieran dos ambientes separados, en cada matero. A cada lado, se sembraron semillas de una de las especies en estudio y se siguió el proceso del tratamiento 1. Tratamiento 4. Plantas en crecimiento o sin competencia. Se tomaron 10 materos; en cinco materos, se sembraron semillas de una especie, en los otros cinco, se plantaron las semillas de la segunda especie y se siguió el procedimiento del tratamiento 1.
Variables evaluadas. Para las dos especies, se midieron las variables peso seco total, peso seco de la parte aérea y peso seco de la parte subterránea, mientras que la producción de inflorescencias solamente se midió en S. vulgaris. Para la medición del peso fresco y el peso seco, se utilizó una balanza digital (Ohaus), con precisión de 0,001g. El peso seco de plantas fue obtenido después de someter las muestras a estufa, hasta llevarlas a peso constante, en un proceso de secado, con temperatura controlada a 70°C, en horno de laboratorio marca Memmer.
En cuanto al número de inflorescencias producidas por S. vulgaris, esta medición inició cuando la planta llegó a madurez fisiológica. Se tomaron los datos de número promedio de semillas producidas en cada inflorescencia y por cada tratamiento.
La recolección de los datos, se inició en el momento en que las plantas de S. vulgaris empezaron la etapa de floración; la recolección de inflorescencias, se realizó cada dos días hasta llegar al final del ciclo de la planta invasora. Al término de este ciclo, se dio por finalizado el ensayo y se recolectaron las plantas, para la toma de los datos de peso fresco foliar, peso fresco radicular y biomasa.
Los datos recolectados fueron analizados separadamente en cada especie. En cuanto al análisis estadístico, se aplicó la prueba no paramétrica de Kruskall-Wallis (Gómez et al. 2003), para validación de hipótesis; respecto al análisis del rendimiento agronómico y el efecto de cada uno de los factores en la competencia, se utilizó la prueba de análisis diferencial en productividad Yield Gap Analysis. Esta metodología, se enfoca en buscar las oportunidades de mejoramiento en cosechas, determinando el efecto de diferentes condiciones de producción sobre el rendimiento agronómico (Ittersum et al. 2013).
RESULTADOS Y DISCUSIÓN
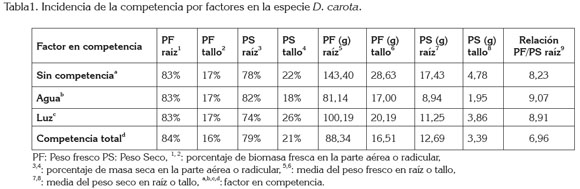
Peso seco y peso fresco de zanahoria: Tal y como se observa en la tabla 1, la masa fresca de la zanahoria, cuando crece sin interferencia alguna o en cualquiera de los factores en competencia, presenta una distribución de sus fotoasimilados, tendiente a mantener un porcentaje de 83%, dirigido a la raíz y 17%, al dosel. El análisis no paramétrico de Kruskall- Wallis arrojó, como resultado, el rechazo de la hipótesis nula, lo que indica la ausencia de diferencias significativas entre esos tratamientos.
Para el caso de peso seco, la variable presenta diferencias significativas entre los tratamientos, según el tipo de competencia, al cual, se estén enfrentando las plantas. Se observa una variación en la distribución de fotoasimilados hacia el dosel y el área radicular. Cuando la planta crece sin competencia, la relación peso seco de raíz y peso seco del tallo, se ubica en 78 y 22%, respectivamente, comportamiento que se observa similar en el caso de la planta al estar bajo condiciones de competencia total, por lo que se infiere, que la relación alométrica raíz:tallo es constante en proporción y que lo que varía es la acumulación total de biomasa en cada órgano, según el tratamiento (Kollmann et al. 2004; Nui et al. 2008). Cuando es sometida a competencia por los factores, luz y agua, se encontró una modificación en el porcentaje de distribución de fotoasimilados; esta relación se modifica a favor de la acumulación en la parte aérea, siendo de un 26%, mientras que un 74% del peso se distribuye hacia la raíz. En el caso de competencia solo por agua, se observa una variación en la translocación hacia la parte aérea -18%-, frente al 74%, de la parte radicular. Este resultado, que puede ser indicador de una respuesta de la planta hacia la competencia en la que se encuentra expuesta y que concordaría con la teoría de equilibrio funcional de Brouwer, según la cual, cuando una planta se encuentra compitiendo por un recurso especifico, cambia el patrón de asignación de los de fotoasimilados, hacia el órgano implicado en la captura del recurso limitante (Camargo & Rodríguez, 2006; Wiener, 2005). En este caso, el recurso limitante es el agua, por lo tanto, es posible que la raíz alcance mayor volumen para interceptar, de forma más eficiente, la absorción del recurso hídrico del suelo y, a su vez, se disminuye el área transpiratoria, para ahorrar agua dentro del sistema de conducción. Esto demuestra que la planta, bajo competencia de un recurso específico, busca favorecer la toma del mismo, aun cuando sacrifique el crecimiento de alguno de sus órganos (Zeng, 2003).
Con base en estos resultados, se evidencia que la distribución de fotoasimilados a cada órgano tiene una composición porcentual homogénea, no sucediendo así para la acumulación neta de los fotoasimilados, pues la variación en cantidad de peso seco es marcada; mientras en los casos de crecimiento sin competencia y en competencia total, se observa una relación de distribución similar (Tabla 1). En los casos de competencia por luz y agua, se puede advertir cómo esta relación varía, aumentando los porcentajes de distribución hacia la raíz cuando la planta es sometida a competencia por agua; en condiciones de competencia por luz, el porcentaje de distribución se ve aumentado hacia la parte foliar, respuesta que indica una redistribución en la asignación de biomasa a la zona fotosintética, para incrementar el área de captura de radiación solar, aspecto apoyado por los planteamientos de Clark & Bullock (2007).
En trabajos afines y realizados por Villamizar et al. (2012), en Lippia dulcis L. y Khurana & Singh (2000), en Albizia procera L., reportaron que en condiciones de estrés hídrico, las especies modifican la asignación de fotoasimilados aumentándolos hacia la raíz, posiblemente, para incrementar el volumen de pelos absorbentes, que garanticen mayor exploración en el suelo y así asegurar mayor captación del recurso hídrico, aspecto que es corroborado por los datos encontrado en el presente trabajo.
Rendimiento agronómico: En cuanto al rendimiento agronómico de la especie cultivada, cuando la zanahoria se manejó en condiciones de crecimiento sin competencia, se encontró que el rendimiento medio estuvo en 143,4g.planta-1, de peso fresco radical y 28,63g.planta-1, de peso fresco en el dosel (Tabla 2). A pesar de conservar proporciones similares de distribución de fotoasimilados, cuando la planta está en competencia total, frente a la planta creciendo sin competencia, el rendimiento de la especie se ve disminuido de manera significativa, en un nivel inferior a cuando compite por el factor luz, en donde pasa de un peso total de 172,03g a 120,38g.planta-1 (Tabla 1). El mayor nivel de sensibilidad por competencia de un factor se encontró cuando la especie está en condiciones de competencia por agua, ya que el análisis de los datos muestra un mayor porcentaje de distribución de biomasa hacia la raíz, pero una menor producción y almacenamiento total, por lo que se podría suponer que el factor agua el crítico para esta hortaliza.
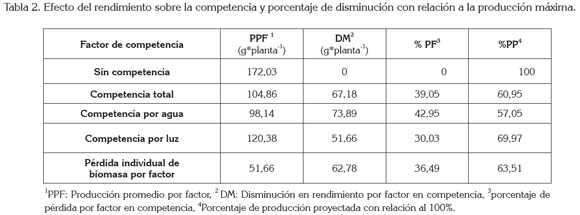
El análisis diferencial de producción aplicado, para determinar el efecto de los factores de competencia sobre las especie (Tabla 2), muestra cómo en condiciones de competencia total un cultivo de zanahoria puede ver reducida la producción, hasta en un 39,05%; cuando el factor en competencia es el agua, se encontraron reducciones del 42,95% y, para el caso de la competencia por luz, la reducción hallada fue del 30,03% (Tabla 2).
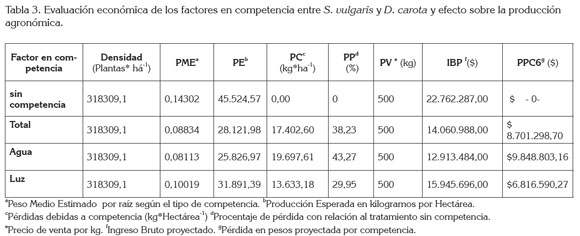
Con base en estos resultados, se modeló un análisis económico, basado en una proyección simulada de los posibles efectos de la competencia, entre S. vulgaris y D. carota, utilizando, como punto de referencia, los siguientes aspectos: un cultivo de una hectárea, una población de 318,309 plantas.ha-1 y un precio de venta, valorado en $500 por kg (DANE, 2012). El análisis reporta que, en condiciones de producción, en los que la planta no compite por recursos, la producción potencial por hectárea estaría en 45,524t (Tabla 3), pero cuando la planta entra a competir por recursos, estos niveles se ven altamente afectados, arrojando pérdidas agronómicas significativas, las cuales, indicarían la necesidad de un adecuado manejo de las plantas invasoras asociadas al cultivo, debido a la alta sensibilidad del cultivo a las diferentes condiciones de competencia.
Efecto de la competencia inter-específica de D. carota sobre S. vulgaris: En la tabla 4, se observan los resultados obtenidos al evaluar el efecto de los diferentes factores en competencia para S. vulgaris, los cuales, revelan que cuando la planta crece sin competencia, la translocación de fotoasimilados, hacia la parte aérea y radicular, se hallan en valores del 61 y 39%, respectivamente, con un peso relativo total de 32,0g, de raíz y 50,454g, en la parte foliar.
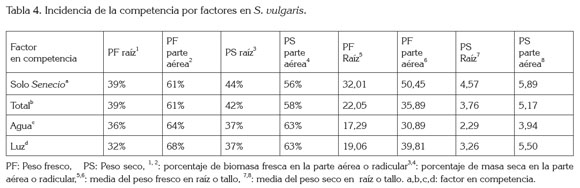
En todos los casos de competencia, los resultados encontrados indican que esta especie realiza una translocación mayor de asimilados hacia el dosel, presentándose una variación del 7%, cuando la planta compite por luz y agua, a favor de la parte foliar. Para el caso de competencia total, se halló, nuevamente, una modificación en el patrón de traslocación de 2%, a favor del dosel. Los porcentajes hallados en peso relativo muestran variación en distribución de fotoasimilados, según el tipo de competencia y esta variación, en la asignación en el caso de competencia por agua, es menor a la presentada, cuando la especie compite por luminosidad, modificándose en 3%, a favor de dosel en competencia por agua y 7%, en competencia por luz.
Los porcentajes de distribución de biomasa en el peso fresco y peso seco en los diferentes tratamientos arrojan una variación en la distribución, según el tipo de competencia a la que se encuentre expuesta la especie, observándose un cambio en los porcentajes de distribución de fotoasimilados, cuando la planta entra en competencia, pero que al ser revisados estadísticamente, arrojan la retención de la hipótesis nula.
En el caso del peso total, la hipótesis nula es rechazada y se puede notar una pérdida general de la producción de biomasa, cuando la especie esta en competencia, en relación a la planta en condiciones ideales de crecimiento. Esta pérdida general de biomasa, sin modificaciones significativas en la asignación de fotoasimilados, posiblemente, sería un indicador que la especie, en condiciones de competencia por recursos, presenta un ajuste en el crecimiento de ella, lo que generaría, como resultado, una menor producción total de biomasa, sin la modificación en los patrones de asignación de los recursos producción, por lo que no cumpliría los postulados de Brouwer, sobre el cambio en la asignación de biomasa, tal y como lo expresan Lei et al. (2013), Jongejans et al. (2006) y Van et al. (1999).
Floración de S. vulgaris: La figura 1 muestra la producción de inflorescencias de S. vulgaris, con una cantidad diferenciada, según el tipo de competencia, al cual, se encuentra expuesta la planta, presentándose una mayor producción de inflorescencias, cuando la planta compite por luz, seguida por la cantidad de flores producidas, cuando la especie no se encuentra en competencia; las menores producciones de flor, se observaron en su orden, cuando la planta compite por agua y cuando se encuentra en competencia total. El análisis de Kruskal-Wallis retiene la hipótesis nula frente a la producción de inflorescencias en cada una de las condiciones de crecimiento, retención que podría indicar una prevalencia de la producción de estructuras reproductivas, a pesar de las condiciones limitantes y que concordaría con el hábito de crecimiento de las plantas invasoras, planteamiento muy similar al reportado por Muller et al. (2000).
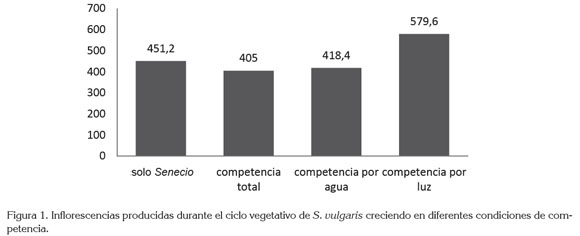
En el caso de la emisión de inflorescencias, se encontró que esta se adelanta cerca de cuatro días, cuando la especie compite por luz, con relación a la planta sin competencia; la velocidad emisión de los botones florales también es más rápida, ya que en 43 días ha emitido el 90% del total de la floración, mientras que cuando la planta no se encuentra en competencia, para los mismos días, apenas ha emitido alrededor del 68% del total de la floración; modificación en la emisión de flores, que ya ha sido notada en especies cuando son sometidas a diferentes condiciones de crecimiento, en las cuales, se encuentra una respuesta que se podría catalogar de naturaleza plástica, afectada por la presión ambiental de un recurso faltante, tal y como lo describen Miner et al. (2005) y Pigliucci (2005).
Finalmente, se observa cómo en el caso de D. carota la existencia de un proceso de competencia genera pérdidas significativas en la producción de biomasa, que en términos de rendimiento agronómico, son importantes en la rentabilidad de la producción.
El comportamiento fisiológico de la especie indicó una respuesta diferencial en la partición de fotoasimilados en la planta y su relación raíz/dosel, siguiendo la teoría expresada por Brouwer (Zeng, 2003), sobre del equilibrio funcional, la cual, indicaría un cambio adaptativo por los recursos limitantes.
Bajo condiciones de competencia por recursos evaluadas, la especie S. vulgaris, mostró una alta producción de estructuras reproductivas, en todos los factores evaluados y mayor, cuando la especie compite por luz; sin embargo, la diferencia en la producción total de biomasa, para cada factor evaluado, el inicio temprano de la fase reproductiva en la planta y el conservar un mismo patrón de translocación y de producción de flor, podría ser un indicador del efecto de la condición ambiental sobre la especie, que podría estar ligado a un ajuste en la velocidad de crecimiento que, a su vez, implica un retraso en los procesos de la planta, sin una modificación en los patrones de asignación de biomasa.
Como conclusión, se deduce que, con base en los resultados encontrados en el presente trabajo, para ambas especies, el efecto individual del factor de competencia más significativo fue la disponibilidad de agua. Ante este efecto, S. vulgaris incrementa la producción de estructuras reproductivas, alcanza un mayor número de inflorescencias y, por tanto, mayor número de semillas, lo que, posiblemente, ocasiona un incremento paulatino en el banco de semillas en el suelo y de ahí la importancia que la especie tiene, en la actualidad, como planta nociva en los cultivos de zanahoria. D. carota disminuye la cantidad de fotoasimilados, para el llenado de raíz, aun cuando, proporcionalmente, se refleja mayor porcentaje de ellos en la relación alométrica raíz:follaje. Esto indica que la planta también ve afectado el desarrollo foliar, por tanto, existirá un inconveniente metabólico, traducido en menores tasas de fotoasimilados, para el llenado final del órgano demanda.
Conflicto de intereses: El manuscrito fue preparado y revisado con la participación de todos los autores, quienes declaramos que no existe conflicto de intereses que ponga en riesgo la validez de los resultados presentados.
BIBLIOGRAFÍA
1. ACOSTA, L.; AGÜERO, R. 2001. El banco de propagulos de malezas enel agroecosistema, conocimiento actual y propuesta metodológica para su estudio. Agr. Mesoam. 12(2):141-151.
2. AULD, B.A. 1996. Criterios económicos para el desarrollo del manejo de malezas. En: Labrada, R.; Caseley, J.C.; PARKER, C. (eds.). Manejo de malezas para países en desarrollo. Estudio FAO Producción y Protección Vegetal, FAO. (Roma). p.253-262.
3. BEGON, M.; HARPER, J.; TOWNSEND, C.R. 1995. Ecology: Individuals, Populations and Communities.Editorial Omega (Barcelona). 1049p.
4. CAMARGO, I.; RODRÍGUEZ, N. 2006. Nuevas perspectivas para el estudio de la asignación de biomasa y su relación con el funcionamiento de plantas en ecosistemas neotropicales. Acta Biol. Col. 11:75-87.
5. CAMBRÓN, V.; SUZÁN, H.; VARGAS, J.; SÁNCHEZ, N.; SÁENZ, C. 2013. Growth strategies and biomass distribution in Pinus pseudostrobus under different conditions of competition. Fitotecnia Mex. 36(1):71-79.
6. CLARK, B.; BULLOCK, S. 2007. Shedding light on plant competition: modelling the influence of plant morphology on light capture (and vice versa). J. Theoret. Biol. 244(2):208-217.
7. DANE. 2012. Encuesta Nacional Agropecuaria. Bogotá. 450p.
8. DOLL, J. 1989. Información básica sobre competencia entre cultivos y malezas.Ed. CIAT(Cali). 42p.
9. GÓMEZ, M.; DANGLOT, C.; VEGA, L. 2003. Sinopsis de pruebas estadisticas no parametricas. Cuando usarlas. Rev. Mex. Pediatria. 70(2):91-99.
10. HÅKANSSON, S. 2003. Weeds and Weed Management on Arable Land: An Ecological Approach. CABI Publishing (Cambridge). 288p.
11. IJLAL, Z.; TANVEER, M.; SAFDAR, A.; AZIZ, M.; ASHRAF, F.; ATIF, A.; MAQBOOL, M. 2011. Effects of weed crop competition period on weeds and yield and yield components of sesame (Sesamum indicum L.). Pakistan J. Weed Sci. Res. 17(1):51-63.
12. ITTERSUM, M.; CASSMAN, K.; GRASSINI, P.; WOLF, J.; TITTONELL, P.; HOCHMAN, Z. 2013. Yield gap analysis—rationale, methods and applications— Introduction to the Special Issue. Field Crops Res. 143:1-3.
13. JANECEK, S.; PATÁCOVÁ, E.; KLIMESOVÁ, J. 2014. Effects of fertilization and competition on plant biomass allocation and internal resources: does Plantago lanceolata follow the rules of economic theory? Folia Geobot. Academic Search Premier. 49(1):49-64.
14. JONGEJANS, E.; SHEPPARD, W.; SHEA, K. 2006. What controls the population dynamics of the invasive thistle Carduus nutans in its native range. J. Appl. Ecol. 43 (5):877-886.
15. KOLLMANN, J.; DIETZ, H.; EDWARDS, P. 2004. Allocation, plasticity and allometry. Perspect. Plant Ecol., Evolut. Syst. 6(4):205-206.
16. KHURANA, E.; SINGH, J.S. 2000. Influence of seed size on seedling growth of Albizia procera under different soil water levels. Ann. Bot. 86(6):1185-1192
17. LEI, L.E.; WEINER, J.; ZHOU, D.; HUANG, Y.; SHENG, L. 2013. Initian density sffects biomass-density and allometric relationships in self-thinning populations of Fagopyrum esculentum. J. Ecology. 101(2):475-483.
18. LOBELL, D.; CASSMAN, G.; FIELD, C. 2009. Crop yield gaps: their importance, magnitudes, and causes. Ann. Rev. Environm. Resources. 34:2-26.
19. McCONNAUGHAY, K.D.; COLEMAN, J.S. 1999. Biomass allocation in plants: ontogeny or optimality? A test along three resources gradients. Ecology. 80:2581-2593
20. McGINLEY, M. 2012. Exploitative competition. Encyclopedy of the Earth. 11p. Disponible desde Internet en http://www.eoearth.org/article/Exploitative_competition&eiCnXvUOfxIozs8 (con acceso 12/7/2013).
21. MINER, B.; SULTAN, S.; MORGAN, S.; PADILLA, K.; RELYEA, A. 2005. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol. 20(12):685-692.
22. MÜLLER, I.; SCHMID, B.; WEINER, J. 2000. The effect of nutrient avaliability on biomass allocation patterns in 27 species of herbaceous plants. Perspect. Plant. Ecol., Evol. System. 3:115-127.
23. NUI, K.; LUO, J.; CHOLER, P.; DU, Z. 2008. The role of biomass allocation strategy in diversity loss due to fertilization. Basic Appl. Ecol. 9(5):485-493.
24. PERRY, J.; PERRY, R. 1989. Interspecific competition between seeds: relative planting date and density affect seedling. Ecology. 70(6):1639-1644.
25. PIGLIUCCI, M. 2005. Evolution of phenotypic plasticity: Where are we going now. Trends Ecol. Evol. 20(9):481-486.
26. PLAZA, G.; QUINTANA, D.; APONTE, L.; CHAVES, B. 2009. Caracterización de la comunidad de malezas en un sistema de producción de rosa bajo invernadero en la Sabana de Bogotá. Agr. Col. 27(3):385-394.
27. QASEM, J.R.; HILL, T.R. 1989. Possible rôle of allelopathy in competition between tomato, Senecio vulgaris L. and Chenopodium album L. Weed Res. 29(5):349-356.
28. RADOSEVICH, S.; HOLT, J.; GHERSA, C. 2007. Ecology of Weeds and Invasive Plants: Relationship to Agriculture and natural resourse management. United States of America: Wiley-intercience Hoboken, NJ, 130p.
29. ROBERTS, H.; POPAI, A. 1970. Ecology of Capsella bursa pastoris (L.) Medik. and Senecio vulgaris L. in relation to germination. J. Ecol. 58(1):123-139.
30. TERRAZAS, F.; GERMAIN, N. 1995. Control de Spergula arvensis L. en cultivo de Trigo. Rev. Agric. 50(24):41- 46.
31. VAN, T.K.; WHEELER, G.S.; CENTER, T.D. 1999. Competition between Hydrilla verticillata and Vallisneria americana as influenced by soil fertility. Aquatic Bot. 62(4):225-233.
32. VILLAMIZAR, M.; RODRÍGUEZ, N.; FERNÁNDEZ, W. 2012. Plasticidad fenotípica en plantas de Lippia dulcis (Verbenaceae) sometidas a déficit hídrico. Acta Biol. Col. 17(2):363-378.
33. VIBRANS, H. 2009. Malezas de México. Disponible desde Internet en: http://www.conabio.gob.mx/malezasdemexico/asteraceae/senecio-vulgaris/fichas/ficha.htm (con acceso 12/2/2014).
34. WIENER, J. 2005. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol., Evolut. System. 6(4):207-215.
35. ZENG, B. 2003. Funtional equilibrium between photosynthetic an above-ground nonphotosynthetic structures of plants: Evidence from a prunning experiment with three subtropical tree species. Acta Bot. Sinica. 45(2):152-157.
36. ZIMDAHL, R.L. 2007. Fundamentals of Weed Science. Estados Unidos: Elsevier. 688p.
Recibido: Junio 3 de 2014 Aceptado: Marzo 4 de 2015

Revista U.D.C.A Actualidad & Divulgación Científica por Universidad de Ciencias Aplicadas y Ambientales se distribuye bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.