CIENCIAS AGROPECUARIAS Y BIOLÓGICAS - Artículo Cient?fico
ANÁLISIS BROMATOLÓGICO EN FRUTOS DE TOMATE (Lycopersicon esculentum Mill) AFECTADOS POR SALINIDAD POR NaCl
BROMATOLOGICAL ANALYSIS IN TOMATO FRUITS (Lycopersicon esculentum Mill) AFFECTED BY NaCl SALINITY
Fánor Casierra-Posada1; César A. Pachón2; Roger C. Ni?o-Medina.3
1Ingeniero Agrónomo PhD., profesor asociado en la Facultad de Ciencias Agropecuarias de la Universidad Pedagógica y Tecnológica de Colombia. Apartado aéreo 661, Tunja ? Boyacá / Colombia. Grupo de Investigación Ecofisiología Vegetal. Correo electrónico:Fanor.Casierra@uptc.edu.co (Autor para correspondencia)
2Ingeniero agrónomo, Universidad Pedagógica y Tecnológica de Colombia, Tunja ? Boyacá. Correo electrónico:capaingagro@latinmail.com
3Estudiante de Ingeniería Agronómica en la Facultad de Ciencias Agropecuarias de la Universidad Pedagógica y Tecnológica de Colombia. Correo electrónico:camilo12355@yahoo.com
Rev. U.D.C.A Act. & Div. Cient. 10 (2): 95-104, 2007
RESUMEN
El estrés salino es uno de los problemas ambientales más serios que limita la productividad en plantas cultivadas. En sistemas de producción bajo invernadero causa problemas osmóticos a las raíces, debido al suministro de agua y de fertilizantes a través de la fertirrigación, con su consecuente acumulación de sales entorno a la zona de las raíces. El propósito de esta investigación fue la evaluación de los efectos de la salinidad por cloruro de sodio (NaCl) sobre algunas características cualitativas de los frutos en cuatro híbridos de tomate (Lycopersicon esculentum Mill). El estudio, se realizó bajo invernadero en Tunja/Colombia en plántulas de los híbridos Supermagnate F1, Marimba F1, Gloria F1 y Astona F1 que crecían en materas con suelo, con capacidad para 3,5kg, se expusieron a 40 y 80mM de NaCl. Las plantas control no fueron tratadas; ésto correspondió a valores de conductividad eléctrica de 5,2 y 8,8dS.m-1, respectivamente y 1,8dS.m-1 para los controles. Las materas, se regaron dos veces por semana y se evitó el exceso de agua. Los resultados mostraron que el porcentaje de proteína, de fibra, de grasas y de carbohidratos fue más elevado en frutos de plantas tratadas con NaCl que en los de las plantas control, mientras que los contenidos de cenizas y de humedad, se redujeron con la salinidad. No se encontraron diferencias estadísticas entre híbridos ni entre la interacción híbridos x salinidad.
Palabras clave:Estrés osmótico, Supermagnate F1, Marimba F1, Gloria F1, Astona F1.
SUMMARY
Salt stress is one of the most serious environmental problems limiting productivity of crop plants. Cropping system under greenhouse conditions causes osmotic problems to roots due to water and fertilizers supplied through fertirrigation, with a consequent salt accumulation around the root-zone. The aim of the present research was the evaluation of the effects of NaCl-salinity on some fruit quality characteristics of four tomato (Lycopersicon esculentum Mill) hybrids. The study was carried out under greenhouse conditions in Tunja/Colombia. Plantlets of hybrids Supermagnate F1, Marimba F1, Gloria F1 and Astona F1 growing on 3.5kg pots filled with soil were exposed to 40 or 8 mM NaCI. Control plants were untreated. These corresponded to electrical conductivities of 5.2 or 8.8 dS.m-1 respectively and 1.8dS.m-1 for controls. Pots were irrigated twice a week and the water excess was avoided. Results showed that percentage of protein, fibre, fat and carbohydrates were higher in fruits of NaCl-treated plants than in those of control plants, while ash and water contents were reduced by salinity. There were not statistical differences between the hybrids nor within the interaction hybrid x salinity.
Key words:Osmotic stress, Supermagnate F1, Marimba F1, Gloria F1, Astona F1.
INTRODUCCIÓN
Alrededor de 10 millones de hectáreas de terrenos cultivables se pierden cada a?o en el mundo a causa de la excesiva salinidad. A lo largo del siglo XX, los científicos habían hecho grandes esfuerzos por obtener plantas de cultivo tolerantes a las sales, por los métodos tradicionales de selección, sin ningún éxito (El País, 2001). Sin embargo, en los últimos a?os, se reportó la obtención exitosa de plantas transgénicas de tomate que toleran la salinidad mediante un mecanismo que les permite acumular los iones tóxicos en las hojas y no en los frutos (Zhang & Blumwald, 2001).
En Colombia, los suelos susceptibles a la salinización cubren una extensión de 86.592km2, de los cuales, un área correspondiente a 90,39%, se encuentra ubicada en zonas secas. Las zonas susceptibles a la salinización abarcan gran parte de la región Caribe, de los valles interandinos (ríos Magdalena y Cauca) y de los altiplanos, donde se desarrolla actualmente y se tiene proyectado ampliar la producción agrícola intensiva del país (Ministerio de Ambiente, Vivienda y Desarrollo Territorial de Colombia, 2004).
El cultivo de tomate reviste gran importancia en todo el mundo (Rick, 1978) y el hecho de que en Colombia esta hortaliza haya cubierto 15.160ha en 2003, con una producción de 392.213 ton (Asohofrucol, 2003) es un índice de la relevancia del cultivo para los agricultores y consumidores colombianos.
Según el sistema de producción de tomate, el cultivo se puede establecer a campo abierto o bajo invernadero, de los cuales, el cultivo bajo cobertura ha generado problemas de salinidad en el sustrato, dado que el riego localizado y la implementación de fertirrigación son causales de salinización de los suelos (Assouline et al. 2006). Se ha encontrado que el uso de aguas de riego con conductividad eléctrica (CE) de 1,7; 2,3; 3,4 y 5,0 reduce en un 0; 10; 25 y 50%, respectivamente, la producción en plantas de tomate (Ayers, 1997). Con base en la CE, la salinidad máxima tolerada por las plantas de tomate es de 2,5dS.m-1, con una reducción del orden de 9,9% por cada unidad en que se incremente la CE, por sobre ese límite (Maas & Hoffman, 1997). Sin embargo, existen reportes que el incremento unitario por sobre 1dS.m-1 reduce la producción de tomate alrededor de 11% (Campos et al. 2006). De esta manera se reporta al tomate como un cultivo moderadamente tolerante a la salinidad (Mass, 1986).
Bajo condiciones normales de cultivo, los frutos de tomate contienen en promedio 94% de agua, 1% de proteína cruda y 0,6% de minerales, expresados como cenizas (Herrmann, 2001). Sin embargo, estos contenidos se modifican si las plantas se ven expuestas a condiciones de estrés osmótico (Adams, 1991).
Existen algunos reportes en los que se indica que el tomate rojo, al igual que muchos otros cultivos hortícolas, tienen un alto requerimiento de potasio en los frutos, para lograr alta calidad (Adams, 1994; Bugarín-Montoya et al. 2002). Sin embargo, el criterio de calidad ha sido motivo de controversia, pues se define con base en características organolépticas determinadas por el genotipo (Poysa, 1992), el manejo del agua (Mitchel & Shennan, 1991), la salinidad (Adams, 1991; Li et al. 2001), el estado nutricional de la planta por el entorno en la raíz y en la parte aérea (Ho & Adams, 1995) y por el manejo poscosecha (Kader et al. 1978). Incluso, se ha documentado que el rendimiento es independiente de la calidad de los frutos cosechados (Ho, 1996).
El efecto de la salinidad sobre el tomate puede ser negativo, debido a la reducción de la producción o el incremento del secamiento apical del fruto (blossom-end rot), causado por problemas nutricionales relacionados con el calcio, conocido entre los cultivadores locales como ?culillo?. Puede tener también un efecto positivo por el incremento en el contenido de sólidos solubles totales (SST) (Mizrahi & Pasternak, 1985; Cuartero & Mu?oz, 1999; De Pascale et al. 2001) y la acidez de los frutos (De Pascale et al. 2001), lo que redunda en un beneficio, cuando los frutos tienen fines industriales.
El objetivo del presente estudio fue la determinación de los cambios inducidos por niveles crecientes de salinidad, sobre las características bromatológicas en frutos de cuatro híbridos de tomate, mantenidos bajo condiciones de invernadero.
MATERIALES Y MÉTODOS
El ensayo se realizó bajo cobertura, en las instalaciones de la Universidad Pedagógica y Tecnológica de Colombia, en Tunja/Colombia, con temperatura promedio de 15,8°C y humedad relativa promedio de de 72%. Para efecto del experimento, se tomaron plántulas de cuatro semanas de edad, que habían germinado en turba. El material vegetal correspondió a plantas de los híbridos Supermagnate F1, Marimba F1, Gloria F1 y Astona F1, proporcionados por Impulsemillas®, las cuales se sembraron en materas con suelo, con capacidad para 3,5kg.
Una vez las plantas reiniciaron el crecimiento y se aplicó al suelo de cada matera una mezcla de fertilizante comercial de alta solubilidad (valores en g.kg-1), así: N: 0,4; P2O5: 0,03; K2O: 0,05; CaO: 0,0005; MgO: 0,0013; S: 0,00137; B: 0,0002; Cu: 0,00014; Fe: 0,00012; Mn: 0,0013; Mo: 0,00005 y Zn: 0,0002.
Los tratamientos para inducir la salinidad con NaCl fueron 0, 40 y 80 mM. kg-1 de suelo seco al aire, los cuales indujeron valores de conductividad eléctrica de 1,8; 5,2; y 8,8dS.m-1, respectivamente. Los tratamientos se aplicaron aproximadamente cuatro semanas después del transplante. La cantidad total de NaCl, se aplicó gradualmente a cada matera a lo largo de un periodo cercano a cuatro semanas. Durante el ensayo, la humedad del suelo de las materas se mantuvo cercana a la capacidad de campo. La cantidad de agua diaria para regar las materas, se calculó con base en el peso de la matera y en la pérdida de agua con respecto al peso de la misma el día anterior, de manera que el suelo en las materas no llegara al punto de marchitez temporal ni sobrepasara la capacidad de campo.
Los frutos se cosecharon ocho semanas después de terminar los tratamientos con la sal, momento en el que se determinaron los contenidos de proteína (Método Kjeldahl), cenizas (Incineración a 600°C durante dos horas), grasas (disolución en éter de petróleo), fibra cruda (secado de muestras a 105°C luego de la adición de KOH 0,556 N y acetona), carbohidratos (por diferencia entre la sumatoria de las cenizas, grasas, fibra y proteína, con el total de la masa seca) y humedad (secado de muestras a 105°C hasta peso constante) (Ranganna, 1977).
El experimento consistió en un ensayo bifactorial, con tres niveles de salinidad y cuatro híbridos, con dise?o completamente al azar y cuatro replicaciones, tomando tres plantas como unidad experimental. La información, se analizó mediante un análisis de variancia y la prueba de separación de promedios de Tukey (P<0,01) con el programa SPSS versión 11.5.
RESULTADOS Y DISCUSIÓN
Como resultado de los análisis estadísticos de la información, no se encontraron diferencias estadísticamente significativas para el factor híbrido ni para la interacción híbrido x salinidad, resultados que coinciden con lo encontrado por Mizrahi (1982), quien evaluó la maduración en frutos de tres cultivares de tomate expuestos a salinidad por NaCl. Sólo hubo diferencia estadísticamente significativa para el factor salinidad, por tanto, los resultados reportados corresponden al promedio de los frutos de los cuatro híbridos evaluados.
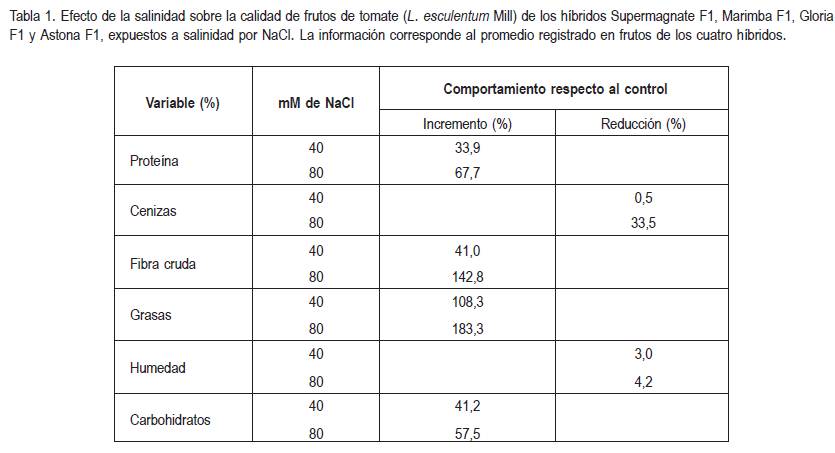
Se encontró un comportamiento diferencial de los factores estudiados, en cuanto a su concentración, con relación al valor registrado para los frutos de las plantas control, cuando las plantas fueron o no expuestas a la salinidad. Es así, como la concentración de NaCl motivó una reducción en los contenidos de humedad y de cenizas en los frutos, mientras que la cantidad de proteína, de fibra cruda, de grasas y de carbohidratos, se vio incrementada como consecuencia de la exposición a las condiciones salinas (tabla 1).
La salinidad indujo un incremento en el contenido de proteína de los frutos, variable en la que se encontraron diferencias altamente significativas, según la prueba de separación de promedios de Tukey (P<0,01). El incremento fue proporcional a la concentración de NaCl en el sustrato, el contenido de proteína aumentó alrededor de 34% con cada incremento de 40mM de NaCl en el suelo (tabla 1, figura 1).
En lo relacionado con el contenido de cenizas, no se encontró diferencias significativas para ninguno de los factores evaluados (híbrido, salinidad y su interacción), a pesar de lo cual, se observó una reducción de 33,5% en frutos de plantas expuestas a 80mM de NaCl, con relación a los frutos de las plantas control, mientras que una contenido de 40mM de NaCl prácticamente no alteró el valor de esta variable, con relación a los controles (tabla 1).
De manera similar a los resultados encontrados en el presente ensayo, se observó un incremento en el contenido de proteínas en frutos de Fragaria sp, como consecuencia de la exposición de las plantas a NaCl en concentraciones de 40 u 80meq.L-1. Sin embargo, los dos cultivares de fresa expuestos a la sal mostraron diferente respuesta en cuanto a la cantidad de proteína registrada en frutos (Keutgen & Keutgen, 2003). Esto puede indicar que el componente genético es determinante en la respuesta de los cultivares, a pesar de que en el presente estudio no se registraron diferencias estadísticamente significativas en cuanto al contenido de proteína, en los híbridos evaluados.
Se ha reportado que bajo condiciones de estrés salino, se acumulan cierto tipo de proteínas llamadas osmotinas, cuya función está aún por definir y que forman cerca de 12% del contenido total de la proteína del tejido. Específicamente, en plantas de tomate, se ha encontrado la osmotina-26, como respuesta al estrés salino por NaCl (Singh et al. 1989). Se requieren condiciones salinas y potencial osmótico alto, para la expresión de los genes que inducen la producción y la acumulación de las osmotinas (La Rosa et al. 1992). Adicionalmente, se ha documentado suficientemente la producción de prolina, extensinas, glicina, proteínas ricas en arabinogalacturano, las cuales se fijan a las paredes celulares bajo condiciones de estrés salino (Showalter, 1993). El contenido de extensinas y de celulosa, en plantas expuestas a la salinidad es una condición de adaptación y de hecho; se ha encontrado un mayor contenido de extensinas y celulosa en paredes celulares de plantas adaptadas a la salinidad, que en aquellas no adaptadas (Iraki et al. 1989), tal como se discutirá con el incremento en el contenido de fibra cruda encontrada en el presente trabajo.
A pesar de esto, la salinidad no siempre representa un factor determinante del incremento en el contenido de proteína en plantas, pues se encontró que en Brassica sp., no hubo diferencia significativa en el contenido de proteína en cultivares expuestos a condiciones salinas (Francois, 1994).
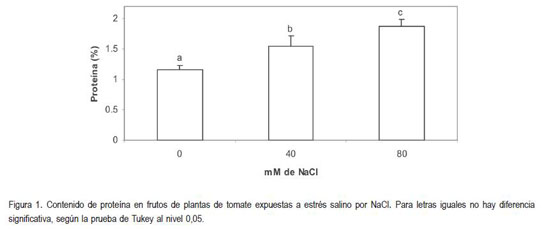
La cantidad de fibra cruda encontrada en los frutos, de acuerdo con las características del ensayo, mostró un incremento directamente proporcional al contenido de NaCl en el sustrato. Al respecto, se encontró diferencia altamente significativa (P<0,01) (figura 2).
Bajo condiciones normales de cultivo, los frutos de tomate pueden contener 0,58g de celulosa, 0,13g de hemicelulosa y 0,26g de lignina, por 100g de materia seca (Herranz et al. 1981). Por otro lado, como el contenido total de fibra en estos tomates estaría en el rango de 1,0 a 1,5% de la materia fresca (Herrmann, 2001). Al igual que en el presente ensayo, se han evidenciado cambios en el contenido de fibra, como consecuencia de la exposición a las sales. Se ha encontrado que la salinidad incrementa la cantidad de fibra y de cristales de oxalato de calcio en la corteza de árboles de casta?a (Aesculus hippocastanum) (Eckstein et al. 1976); además, en plantas de Salix sp., se encontró que luego de la exposición a la salinidad, el tejido xilemático contenía menos vasos y más fibra en individuos tolerantes a las sales, que en aquellos sensibles (Eckstein et al. 1978). Además, se ha encontrado que, con frecuencia, la salinidad promueve la suberización de la hipodermis y la endodermis de las raíces con la formación de una banda de Caspari más cerrada en el ápice de la raíz que en raíces no expuestas a la salinidad (Walker et al., 1984). De igual manera, las paredes celulares en plantas expuestas a la salinidad, se vuelven densas y onduladas (Shannon et al. 1994).
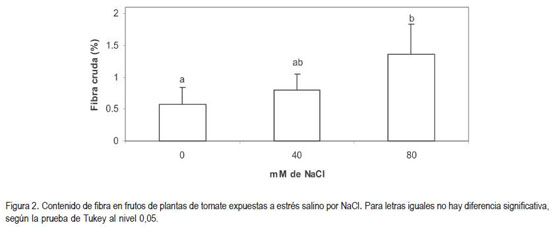
Los resultados revelaron un incremento en el porcentaje de grasas en los frutos de plantas sometidas a las condiciones salinas. El aumento en esta variable fue directamente proporcional a la cantidad de NaCl adicionada al suelo en el que crecían las plantas. Al respecto, se encontró diferencia altamente significativa (P<0,01) (figura 3).
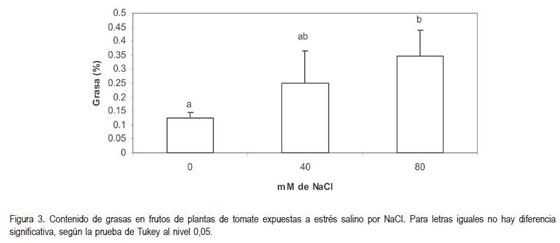
Por el contrario, a los resultados encontrados en este trabajo, el estrés osmótico por NaCl y CaCl2 no incrementó el contenido de aceites en semillas de canola (Brassica sp.) (Francois, 1994), ni en Curthumus tinctorius (Irving et al. 1988). El incremento en el contenido de grasas, como resultado de la salinidad encontrado en los frutos analizados en el presente estudio, puede ser un mecanismo de adaptación que contribuiría a la osmoregulación.
Se encontró diferencia altamente significativa (P<0,01) en el análisis de la información de la variable contenido de humedad en los frutos, entre el testigo y las plantas tratadas con NaCl (figura 4), observándose que bastaba una concentración de 40mM para que el contenido de humedad se redujera en términos de 3%; por sobre esa concentración de sal, la reducción, prácticamente permanece invariable, pues no se encontró diferencia significativa entre el contenido de humedad en frutos de plantas que crecían en un sustrato enriquecido con 40 u 80mM de NaCl.
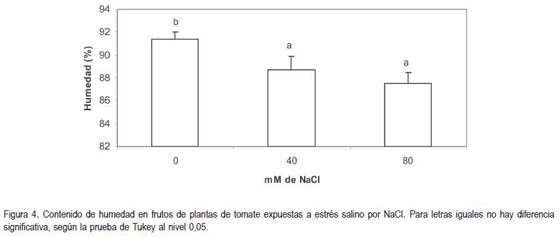
Debido a las propiedades de la membrana y de la pared, la célula puede perder agua con facilidad, pues los cambios en el potencial osmótico de la solución del suelo inducidos por la salinidad, restringen considerablemente la toma de agua; adicionalmente, los tejidos pierden agua mediante la transpiración; por tanto, una de las primeras respuestas de los tejidos a la salinidad, es la pérdida de la turgencia en los tejidos (Levitt, 1980). La turgencia en la planta, se debe mantener, pero así la planta pierde agua por transpiración, además que la toma de agua por las raíces es limitada. Esto conduce a que la disponibilidad de agua para el crecimiento y de elongación de las células de los frutos se vea bastante comprometida, lo cual se manifiesta con una reducción en el contenido de humedad de los frutos, como se pudo observar en este ensayo.
Como consecuencia de la salinidad en el sustrato, se encontró un incremento en el contenido de carbohidratos en los frutos, con relación a los frutos de plantas no tratadas con NaCl (figura 5). El análisis estadístico de esta variable reveló diferencias altamente significativas (P<0,01) entre los frutos de las plantas control y los de aquellas expuestas a la salinidad, pero no hubo diferencia significativa entre los dos niveles de salinidad evaluados (figura 5). Bajo las condiciones en que se desarrolló el ensayo, se debería esperar una reducción en el contenido de carbohidratos en frutos en el rango de 41,2 a 57,5% cuando las plantas de tomate confrontan contenidos de 40 a 80mM de NaCl en el sustrato (tabla 1).
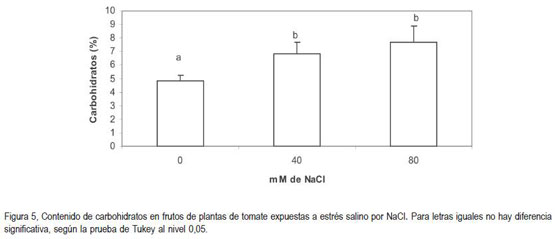
Varios autores coinciden en afirmar que el efecto de la CE del sustrato en el que crecen las plantas de tomate sobre el contenido de materia seca en los frutos es directamente proporcional (Sonneveld & Welles, 1988; D?Amico et al. 2003). Se ha encontrado que el aumento en la CE, como consecuencia de la salinidad, induce un incremento en el contenido de materia seca en los diferentes órganos (Casierra-Posada & García, 2005; Casierra-Posada & Hernández, 2006). Este resultado, se pude justificar mediante el hecho de que las plantas acumulan una cantidad de osmolitos en los tejidos con el propósito de hacer el ajuste osmótico, pero dado el alto poder vertedero de los frutos en desarrollo, éstos productos se acumulan en ellos, produciendo un aumento relativo en el contenido de carbohidratos, dado que se reduce el contenido de humedad en los frutos, lo que también sucede con el contenido de materia seca en los mismos (Casierra-Posada & García, 2006), como se encontró en el presente ensayo y se presenta en las figura 4 y figura 5.
Los tejidos de plantas sometidas a estrés salino usualmente manifiestan una reducción en el contenido de almidón, el cual se reemplaza por un incremento en el contenido de azúcares (Mohan & Gupta, 1997, Sen & Mohammed, 1987; Mohammed et al. 1998). Ésto se pudo evidenciar en los frutos analizados, en los cuales el contenido de carbohidratos se incrementó proporcionalmente a la concentración de NaCl en el sustrato. Los osmolitos producidos pueden, por un lado, contribuir al incremento del potencial osmótico de los tejidos, facilitando la retención de agua en el citoplasma y, por otro lado, pueden cumplir con funciones de protección (Schulze et al. 2005).
Como información adicional vale la pena mencionar que se observó que la salinidad indujo una maduración más rápida de los frutos en comparación con los de las plantas control, aunque no se cuantificó el valor de la reducción en términos de tiempo. Este resultado fue reportado también por Mizrahi (1982), quien encontró una reducción de 4 a 15% en el tiempo de desarrollo de los frutos sometidos a condiciones salinas. Por otro lado, el mismo autor menciona que el sabor de los frutos de plantas sometidas a salinidad era más agradable que el de los de plantas control, lo cual es el resultado de los azúcares acumulados en los frutos, como mecanismo de regulación osmótica. Este aspecto podría ser beneficioso para los consumidores de tomates en fresco, aunque se debe evaluar la relación beneficio costo de la producción de tomates bajo estas condiciones, dado que se observó que la salinidad reduce considerablemente el rendimiento, en términos de cantidad de fruta cosechada y de tama?o de frutos.
AGRADECIMIENTOS
Los autores agradecen la colaboración de Impulsemillas® en el suministro del material vegetal; así como el apoyo del personal del Laboratorio de Análisis de Alimentos de la Escuela de Química de Alimentos de la UPTC, por la realización de los análisis reportados en este texto.
Este estudio, se desarrolló con el apoyo de la Dirección de Investigaciones (DIN) de la Universidad Pedagógica y Tecnológica de Colombia, en el marco del plan de trabajo del grupo de investigación Ecofisiología Vegetal, adscrito al programa de Ingeniería Agronómica de la Facultad de Ciencias Agropecuarias.
BIBLIOGRAFÍA
1. ADAMS, P. 1991. Effects of increasing the salinity of the nutrient solution with major nutrient or sodium chloride on yield, quality and composition of tomatoes grown in rockwool. J. Hortic. Sci. 66:210-217.
2. ADAMS, P. 1994. Nutrition of greenhouse vegetables in NFT and hydroponics system. Acta Hort. 361:245-257.
3. ASOHOFRUCOL. 2003. Tomate. Disponible desde Internet en: http://www.asohofrucol.com.co/archivos/biblioteca/biblioteca_30_GUIAhortifruticultura[1].pdf(con acceso10/08/2007).
4. ASSOULINE, S.; M?LLER, M.; COHEN, S.; BEN-HUR, M.; GRAVA, A.; NARKIS, K.; SILBER, A. 2006. Soil-plant system response to pulsed drip irrigation and salinity: Bell pepper case study. Soil Sci. Soc. Am. J. 70:1556-1568.
5. AYERS, R.S. 1997. Quality of water for irrigation. J. Irrig. Drain. E.-Asce, Division 103:135-154.
6. BUGARÍN-MONTOYA, R.; GALVIS-SPINOLA, A.; SANCHEZ-GARCÍA, P.; GARCÍA-PAREDES, D. 2002. Demanda de potasio del tomate tipo saladette. Terra. 20(4):391-399.
7. CAMPOS, C.A.B.; FERNANDES, P.D.; GHEYI, H.R.; BLANCO, F.F.; GON?ALVES, C.B.; CAMPOS, S.AF. 2006. Yield and fruit quality of industrial tomato under saline irrigation. Sci. Agric. (Piracicaba, Brasil). 63(2):146-152.
8. CASIERRA-POSADA, F.; GARCÍA R., N. 2005. Crecimiento y distribución de materia seca en cultivares de fresa (Fragaria sp.) bajo estrés salino. Agr. Col. 23(1):83-89
9. CASIERRA-POSADA, F.; GARCÍA R., N. 2006. Producción y calidad de fruta en cultivares de fresa (Fragaria sp.) afectados por estrés salino. Rev. Fac. Nal. Agr. Medellín. 59(2):3527-3542.
10. CASIERRA-POSADA, F.; HERNÁNDEZ, H. L. 2006. Evapotranspiración y distribución de materia seca en plantas de mora (Rubus sp.) bajo estrés salino. Rev. U.D.C.A Act. & Div. Cient. 9(1):85-95.
11. CUARTERO, J.; MU?OZ, R.F. 1999. Tomato and salinity. Sci. Hortic.-Amsterdam 78:83-125.
12. D?AMICO, M.L.; IZZO, R.; TOGNONI, F.; PARDOSSI, A.; NAVARI-IZZO, F. 2003. Application of diluted sea water to soilless culture of tomato (Lycopersicon esculentum Mill.): effects on plant growth, yield, fruit quality and antioxidant capacity. Food, Agr. & Environ. 1(2):112-116.
13. DE PASCALE, S.; MAGGIO, A.; FOGLIANO, V.; AMBROSINO, P.; RITIENI, A. 2001. Irrigation with saline water improves carotenoids content and antioxidant activity of tomato. J. Hortic. Sci. Biotech. 76:447-453.
14. ECKSTEIN, D.; LIESE, W.; PARAMESWARAN, N. 1976. On the structural changes in wood and bark of a salt-damaged horse chestnut tree. Holzforschung 30:173-178.
15. ECKSTEIN, D.; LIESE, W.; PLOSSL, J. 1978. Histometrische Untersuchungen zur unterschiedlichen Streusalztoleranz von Weiden (Salix spp.). Forstwiss. Centralbl. 97:335-341.
16. El PAÍS. 2001. Dise?ado un tomate transgénico capaz de crecer en suelo salinizado. Disponible desde Internet en: http://centros.edu.xunta.es/iesmendino/seminarios/seminariobio/010731-tomate.PDF (con acceso 15/10/2005).
17. FRANCOIS, L.E. 1994. Growth, seed yield, and oil content of canola grown under saline conditions. Agron. J. 86:233-237.
18. HERRANZ, J.; VIDAL-VALVERDE, C.; ROJAS-HIDALGO, E. 1981. Cellulose, hemicellulose and lignin content of raw and cooked spanish vegetables. J. Food. Sci. 36:1927-1933.
19. HERRMANN, K. 2001. Inhaltsstoffe von Obst und Gem?se. Eugen Ulmer Verlag. Stuttgart. 200p.
20. HO, L.C. 1996. Tomato. En: Zamsky, E.; Schaffer, A.A. (eds.). Photoassimilate distribution in plants and crops. Marcel Dekker, Nueva York. p.709-728.
21. HO, L.C.; ADAMS, P. 1995. Nutrient uptake and distribution in relation to crop quality. Acta Hort. 396:33-44.
22. IRAKI, N.N.; BRESSAN, R.A.; CARPITA, N.C. 1989. Extracellular polysaccharides and proteins of tobacco cell cultures and changes in composition associated with growing-limiting adaptation to water and saline stress. Plant Physiol. 91: 54-61.
23. IRVING, D.W.; SHANNON, M.C.; BREDA, V.A.; MACKEY, B.E. 1988. Salinity effects on yield and oil quality of high-linoleate and high-oleate cultivars of safflower (Carthamus tinctorius L.). J. Agric. Food Chem. 36:37-42.
24. KADER, A.A.; MORRIS, L.L.; STEVENS M.A.; ALBRIGHT-HORTON, M. 1978. Composition and flavor quality of fresh market tomatoes as influenced by some postharvest procedures. J. Am. Soc. Hort. Sci. 103:6-13.
25. KEUTGEN, A.J.; KEUTGEN, N. 2003. Influence of NaCl salinity stress on fruit quality in strawberry. Acta Hort. 609:155-157.
26. LA ROSA, P.C.; CHEN, Z.; NELSON, D.E.; SINGH, N.K.; HAEGAWA, P.M.; BRESSAN, R.A. 1992. Osmotin gene expression is posttranscriptionally regulated. Plant Physiol. 100:409-415.
27. LEVITT, J. 1980. Responses of plant to environmental stresses. Vol II: Water, radiation, salt and other stresses. Academic Press. p.365-490.
28. LI, L.Y.; STANGHELLINI, C.; CHALLA, H. 2001. Effect of electrical conductivity and transpiration on production of greenhouse tomato. Sci. Hortic.-Amsterdam 88:11-29.
29. MASS, E.V. 1986. Salt tolerance of plants. Appl. Agric. Res. 1(1):12-26.
30. MAAS, E.V.; HOFFMAN, G.J. 1997. Crop salt tolerance ? current assessment. J. Irrig. Drain. E.-Asce, División 103:115-134.
31. MINISTERIO DE AMBIENTE, VIVIENDA Y DESARROLLO TERRITORIAL DE COLOMBIA. 2004. Plan de acción nacional de lucha contra la desertificación y la sequía en Colombia. MAVDT, Dirección de Ecosistemas. 124p.
32. MIZRAHI, Y. 1982. Effect of salinity on tomato fruit ripening. Plant Physiol. 69: 966-970
33. MIZRAHI, Y.; PASTERNAK, D. 1985. Effect of salinity on quality of various agricultural crops. Plant Soil. 89:301-307.
34. MITCHEL, J.R.; SHENNAN, C. 1991. Tomato fruit yield and quality under water deficit and salinity. J. Am. Soc. Hort. Sci. 116:215-221.
35. MOHAMMED, S.; KASERA, P.K.; CHAWAN, D.D.; SEN, D.N. 1998. Eco-physiology of Cassia italica (Mill) Lamk. ex Anders in Indian desert. Sci. Cult. 64:233-234
36. MOHAN, H.W.; GUPTA, P. 1997. Plant life under extreme environments. Curr. Sci. 72:306-315.
37. POYSA, V. 1992. Use of Lycopersicon cheesmanii and L. chemielewskii to increase dry matter content of tomato fruit. Can. J. Soil Sci. 73:273-279.
38. RANGANNA, S. 1977. Manual of analysis of fruits and vegetable products. McGraw-Hill. 634p.
39. RICK, L.M. 1978 The tomato. Sci. Amer. 239:67-76.
40. SEN, D.N.; MOHAMMED, S. 1987. Eco-physiological studies of Fagonia cretica in Indian desert. En. Agarwal, S.K.; Garg, R.K. (eds). Environmental issues and research in India. Himanmshu Publications. p.61-83.
41. SCHULZE, E.D.; BECK, E.; M?LLER-HOHENSTEIN, K. 2005. Plant ecology. Springer Verlag. Heidelberg. p.145-194.
42. SHANNON, M.C.; C.M. GRIEVE; L.E. FRANCOIS. 1994. Whole-plant response to salinity. In Plant--Environment Interactions. Ed. R.E. Wilkinson. Marcel Dekker, New York, pp 199-244.
43. SHOWALTER, A.M. 1993. Structure and function of plant cell wall proteins. Plant Cell. 5:9-23
44. SINGH, N.K.; NELSON, D.E.; KUHN, D.; HASEGAWA, P.M.; BRESSAN, R.A. 1989. Molecular cloning of osmotin and regulation of its expression by ABA and adaptation to low water potential. Plant Physiol. 90:1096-1101.
45. SONNEVELD, C; WELLES, G.W.H. 1988. Yield and quality of rockwool-grown tomatoes as affected by variations in EC-value and climatic conditions. Plant Soil. 111:37-42.
46. WALKER, R.R.; M. SEDGLEY; M.A. BLESING; T.J. DOUGLAS. 1984. Anatomy, ultrastructure and assimilate concentrations of roots of citrus genotypes differing in ability for salt exclusion. J. Exp. Bot. 35:1481-1494.
47. ZHANG, H.X.; BLUMWALD, E. 2001. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 19:765-768.
Recibido: Agosto 17 de 2007; Aceptado: Octubre 16 de 2007

Revista U.D.C.A Actualidad & Divulgación Científica por Universidad de Ciencias Aplicadas y Ambientales se distribuye bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.