CIENCIAS AGROPECUARIAS Y BIOLÓGICAS - Artículo Cient?fico
AMPLITUD DE NICHO Y SELECTIVIDAD ALIMENTARIA DEL BORUGO DE PÁRAMO (Cuniculus taczanowskii) (STOLZMANN 1885) EN UN BOSQUE ANDINO NUBLADO (ZIPACÓN ? CUNDINAMARCA)
NICHE BREATH AND DIETARY SELECTIVITY OF THE MOUNTAIN PACA (Cuniculus taczanowskii) (STOLZMANN 1885) IN AN ANDEAN CLOUD FOREST (ZIPACÓN ? CUNDINAMARCA)
Karin Osbahr1; Jesús David Ortiz Montero2; Jairo Pérez-Torres.3
1LÍder Grupo de Investigación en Fauna Silvestre. Universidad de Ciencias Aplicadas y Ambientales U.D.C.A. Dirección para correspondencia: Calle 222 No. 55-37, Bogotá, D.C. Correo electrónico:kosbahr@udca.edu.co
2Estudiante de Biología Pontificia Universidad Javeriana. Dirección para correspondencia. Correo electrónico:jesus.ortiz@javeriana.edu.co
3Director Laboratorio de Ecología Funcional. Departamento de Biología. Pontificia Universidad Javeriana. Dirección para correspondencia: Cra. 7 No. 43-82 Bogota, D.C. Colombia.
Rev. U.D.C.A Act. & Div. Cient. 10 (2): 105-114, 2007
RESUMEN
Cuniculus taczanowskii es una especie típica de la región andina amenazada por factores, como la fragmentación del paisaje y la cacería. El presente trabajo, se desarrolló con el objetivo de establecer la amplitud de nicho, la selección alimentaria, la relación entre la frecuencia de ocurrencia de los ítems alimentarios recuperados en materia fecal y la abundancia de las especies vegetales de consumo potencial colectadas en un bosque andino nublado. Se identificaron 16 items alimentarios pertenecientes a las plantas recogidas como recurso potencial alimentario. Asplenium sp., Thibaudia floribunda y Phytolacca sp. fueron las especies más consumidas. La amplitud de nicho de la especie animal fue baja y no se encontró una relación significativa entre la abundancia de las especies vegetales colectadas y la frecuencia de los fragmentos epidermales encontrados. La especie discrimina entre los estados fenológicos de los recursos evaluados tendiendo a ser selectiva.
Palabras clave: Análisis microhistológico, amplitud de nicho, bosque altoandino, C. taczanowskii, selección alimentaria.
SUMMARY
Cuniculus taczanowskii is a typical species of the Andean region threatened by factors such as landscape fragmentation and hunting. In this work the niche breadth, the dietary preferences, the relationship between the frequency of the dietary items in fresh feces and the abundance of collected plant species at an Andean cloud forest was studied. 16 dietary items were identified as plants used as food source. Asplenium sp., Thibaudia floribunda and Phytolacca sp. where the most consumed plant species. The niche breadth of the species was low and the relationship found between the abundance of the collected plant species and the frequency of the epidermal fragments was not significant. The species discriminates between the phenological stage of the food resources showing a tendency to be selective.
Key words: Microhistological analysis, niche breath, cloud forest, C. taczanowskii, dietary selectivity.
INTRODUCCIÓN
El ?borugo de páramo? (Cuniculus taczanowskii) es una especie de roedor herbívoro de la familia Cuniculidae, con una distribución en Colombia, que abarca los bosques altoandinos (Osbahr, 1995) y cuya presencia ha sido documentada para el cerro de Manjuí, Zipacón (Cundinamarca) (CAR, 2005). En la actualidad, la zona mencionada ha venido sufriendo una drástica transformación del paisaje y un cambio considerable en el uso de la tierra, debido a la expansión agrícola y ganadera (Etter & Wyngaarden, 2000; CAR, 2005). La pérdida de hábitat derivada de esta modificación del paisaje, junto con la cacería indiscriminada, han contribuido a la disminución de las poblaciones naturales de la especie (Nú?ez & Jorgenson, 1996; Osbahr, 1996a; 1996b). Aún cuando la especie se encuentra categorizada como en bajo riego de amenaza (LR/nt) (Baillie, 1996), se considera que su status actual debe ser revisado IUCN (2007).
La evaluación de los hábitos alimentarios de las especies herbívoras ha estado basada, principalmente, en los análisis de los contenidos estomacales y de la materia fecal (Storr, 1961; Sparks & Malechek, 1968; Reichman, 1975; Tobin et al. 1994; Holechek et al. 1982; Cortés et al. 2002). La principal desventaja del análisis de los contenidos estomacales es que implica el sacrificio de un gran número de animales (Reichman 1975, Tobin et al. 1994). Aún cuando han surgido nuevas técnicas basadas en el análisis fecal, su utilización en campo, junto con las características propias de las especies a evaluar, requiere que sean validadas para cada caso, ya que la digestión y la fragmentación diferencial puede subestimar o sobrestimar los ítems alimentarios identificados en las heces (Batzli & Cole, 1979; Holechek et al. 1982; Bartolomé et al. 1995; Kronfeld & Dayan, 1998; Lee & MacGregor, 2004).
Considerando que la amplitud de nicho se relaciona con la disponibilidad relativa de determinado recurso empleado por una especie, su distribución espacial y por ende con el costo en términos de energía y tiempo que debe invertir para su localización (Dajoz, 2002; Guerrero et al. 2002; Lou & Yurrita, 2005), el entendimiento de la selección trófica es de utilidad para planear reservas naturales, entender las consecuencias de programas de reintroducción de especies (Hartman, 1994; Cole et al. 1995) y para predecir el hábitat potencial en el que se podría desarrollar una especie dada (Drickamer, 1971). Para el caso de C. taczanowskii, la información generada al respecto y teniendo en cuenta los factores relacionados como riqueza de la dieta, accesibilidad a las especies vegetales que hacen parte de la dieta, entre otros, son útiles como elementos para la toma de decisiones en cuanto al dise?o de áreas de reserva y planes de manejo (Pérez-Torres, 2002).
El conocimiento de la dieta de C. taczanowskii, se ha basado en estudios generales para el manejo de la especie en cautiverio (Castro, 1996; Amaya & Rodríguez, 2004) y en reportes de plantas consumidas por la especie en su hábitat natural (Delgado et al. 1991; ECOTECNIA, 1997; Nú?ez & Jorgenson, 1996; Osbahr, 1996a; 1999). Aún cuando se cuenta con adelantos en el conocimiento sobre el uso de hábitat (Nú?ez & Jorgenson, 1996; Sánchez, et al. 1999; Pardo, 2004) y a la distribución del recurso alimenticio en el hábitat boscoso (Osbahr, 2003; 2004), aún no se tiene información acerca de la amplitud de nicho de la especie y a la selección de la dieta con relación al recurso alimenticio disponible. Por esta razón, en el presente estudio se evaluó la dieta del borugo de páramo mediante la técnica microhistológica para determinar la relación entre las preferencias alimentarias encontradas y la abundancia de las plantas disponibles en el hábitat natural.
MATERIALES Y MÉTODOS
El estudio se llevó a cabo en el cerro de Manjuí, Zipacón, área que se localiza en jurisdicción del departamento de Cundinamarca, Colombia (04° 48' 20.64'' N 74° 23' 26.55'' W). El rango altitudinal abarca desde los 2550 m.s.n.m., a los 3200 m.s.n.m. (CAR, 2005), prevaleciendo el clima típico frío con temperaturas inferiores a los 17° C (Parra et al. 1970). La distribución de la precitación es bimodal, adquiriendo sus máximos valores entre los meses de abril y julio; decrece entre agosto y septiembre, con una alta tendencia a ascender en el mes de octubre y noviembre (CAR, 2005).
La vegetación del sitio de estudio ha sido fuertemente transformada por la comunidad adyacente a la zona, generando fragmentos de bosque rodeados de zonas de uso agropecuario. Sin embargo, la vegetación remanente aún conserva elementos propios de la región andina encontrándose, en general, especies como Alnus acuminata, Drymis granatensis, Cassia tomentosa, Oreopanax sp., Clusia multiflora, Asplenium praemorsum, Blechnum occidentale, Pellaea ternifolia, Hesperomeles heterophylla, Vallea stipularis, Juglans sp., Ficus sp., Weinmania sp. y Phyllantus spi. entre otros (CAR, 2005; Parra et al. 1970). En el área de estudio se muestrearon cuatro fragmentos de bosque los cuales fueron denominados como bosque 1 (B1), bosque 2 (B2), bosque 3 (B3) y Bosque 4 (B4).
Las especies vegetales evaluadas, como recurso potencial alimentario, fueron colectadas aplicando la técnica de muestreo preferencial teniendo en cuenta la información proporcionada por los trabajos de Osbahr (1999; 2003). La abundancia de las especies fue determinada mediante el establecimiento de cuatro cuadrantes de 10 x 10 m, en cada uno de los bosques. El material vegetal, se colectó por duplicado conservando las plantas de referencia en la colección biológica de la Universidad de Ciencias Ambientales y Aplicadas U.D.C.A, previo secado en un horno, a una temperatura entre 40-45°C (Gavi?o et al. 1974) y almacenando las muestras para el análisis microhistológico en bolsas Ziploc con el fin de evitar su deshidratación (Storr, 1961; Osbahr, 1999).
Para cada planta colectada, se obtuvieron micropreparados de referencia de tallos, hojas, frutos y rizomas mediante la técnica microhistológica modificada por Osbahr (1999). De cada uno de los fragmentos epidermales identificados para cada especie, se obtuvieron microfotografías con una cámara Canon y un aumento de 10X.
Las muestras fecales, se colectaron en diferentes letrinas realizando recorridos sistemáticos en los cuatro bosques seleccionados para asegurar una representación total de la población. Las muestras, frescas y escogidas siguiendo los criterios establecidos por Nú?ez & Jorgenson (1996), se procesaron de acuerdo a lo sugerido por Osbahr (1999), para obtener cinco micropreparados por muestra. Para cada lámina, se observaron 20 campos microscópicos (aumento 40X) para un total de 100 campos evaluados por muestra (Sparks & Malechek, 1968). Se excluyeron los campos que contenían menos del 50% del área ocupada por estructuras epidérmicas (Meserve, 1981). Cada ítem identificado por campo microscópico fue registrado, contado y microfotografiado y se calculó el índice de selección estandarizado (Bi)) (Manly et al. 1993), para determinar la proporción de especies en la dieta (Oi) del borugo de páramo, con relación a la disponibilidad de los recursos alimentarios evaluados en el hábitat (Pi). Dado que el cálculo del índice de selección puede presentar sesgos de acuerdo al tipo de muestreo, se siguió el método sugerido por Johnson (1980) estableciendo la diferencia entre los rangos del recurso determinado en la dieta (ri) y la disponibilidad en el hábitat (si), para así obtener una medida de preferencia relativa (ti).
La amplitud de nicho de la especie, se calculó teniendo en cuenta que la abundancia de los recursos alimentarios disponibles es variable (Krebs, 1999), por lo cual, se utilizó el índice de Hurlbert (1978), el cual se estandarizó a una escala entre 0 ? 1. La intensidad de la relación entre la abundancia de las especies vegetales colectadas como recurso alimentario y los ítems alimentarios encontrados en las heces, se llevó a cabo mediante la correlación de Pearson (r) (Daniel, 2004).
RESULTADOS Y DISCUSIÓN
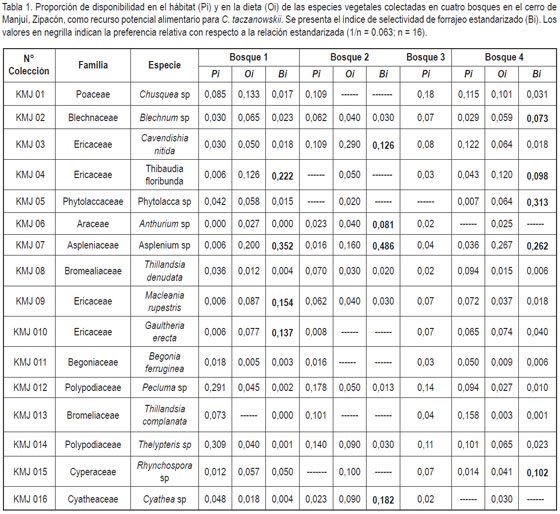
Se colectaron un total de 16 especies vegetales, las cuales fueron consideradas como recurso potencial alimentario para el borugo de páramo (tabla 1). Las especies más abundantes en el área de estudio fueron: Chusquea sp, Tillandsia complanata, Cavendishia nitida al igual que Pecluma sp y Thelypteris sp. (figura 1).

Aún cuando se colectaron un total de 37 muestras de materia fecal, únicamente se utilizaron para el procedimiento microhistológico 20 muestras frescas al cumplir con los requisitos de color y de textura establecidos por Nú?ez & Jorgenson (1996). Cabe anotar que en el bosque 3 no se encontraron muestras de materia fecal, lo cual se puede explicar por la falta de cuevas superficiales y/o de madrigueras apropiadas para que la especie estableciera sus letrinas para la defecación (Osbahr, 1996b). Se identificaron y cuantificaron un total de 2224 fragmentos epidermales, de los cuales 1801 (80,98%) equivalen a tallos, hojas, frutos o rizomas de las 16 especies de plantas verificadas de consumo (tabla 1), mientras que solo 423 fragmentos (19,01%) fueron fragmentos epidermales no reconocidos por no encontrarse en la colección de referencia. De los fragmentos no reconocidos, se debe decir que posiblemente pueden corresponder a partes de una misma planta, por lo que no es proporcional a un número determinado de plantas. De los fragmentos epidermales no reconocidos posiblemente muchos de éstos pertenecen a fibras y tejidos conductores (xilema y floema), los cuales son menos digeribles en comparación con las hojas (Randolph et al. 1991, Cortés et al. 2002). Este hecho puede ser respaldado, ya que los individuos adultos de esta especie consumen grandes cantidades de fibras (Osbahr, 1996a), lo cual se relaciona con la presencia de un ciego grande en su tracto digestivo que facilita la digestión de este tipo de material fibroso (Parra & Osbahr 2000).
Las especies Gaultheria erecta, Pecluma sp., Thelypteris sp. y Rynchospora sp. identificadas en las heces de C. taczanowskii equivalen a plantas que no habían sido reportadas en trabajos previos (Delgado et al. 1991; ECOTECNIA, 1997; Nú?ez & Jorgenson, 1996; Osbahr, 1996a; 1999; 2003).
Las plantas con los mayores porcentajes en la dieta de C. taczanowskii en el cerro de Manjuí, Zipacón fueron: Asplenium sp., Chusquea sp. y Thibaudia floribunda (figura 1). Sin embargo, al calcular globalmente el índice de selección estandarizado, se encontró que C. taczanowskii registró preferencias relativas respecto a Th. floribunda (Bi = 0,188), Phytolacca sp. (Bi = 0,138), Asplenium sp. (Bi = 0,306), mientras que las once plantas restantes registraron valores inferiores a la relación estandarizada (1/n = 0.063; n = 16), por lo cual, se consideran de rechazo relativo (Manly et al. 1993). El valor del índice de selección para Rhynchospora sp. (Bi = 0,063), al ser equivalente a la relación estandarizada, indica que C. taczanowskii rechaza su consumo. La eventualidad que la interpretación del análisis del índice de preferencia se puede sesgar de acuerdo al hecho que un recurso consumido rara vez puede ser abundante en el medio o viceversa (Krebs 1999), se evidenció al aplicar el índice de rangos de preferencia sugerido por Johnson (1980) incluyendo a Chusquea sp. y C. nitida, como especies altamente preferidas por C. taczanowskii modificando, a su vez, las preferencias relativas con respecto a los demás recursos alimentarios evaluados (figura 2).

La dieta evaluada para C. taczanowskii permitió corroborar el consumo tanto de hojas (52,90%), seguido por el rizoma (20,22%), el tallo (14,28%) y los frutos (12,57%). Aún cuando el elevado consumo de hojas se explica por el hecho que se encontraron fragmentos vegetales de las 16 especies de plantas evaluadas, al calcular el índice de selección, se encontró que C. taczanowskii es selectivo en su consumo, con preferencias relativas respecto a Th. floribunda (Bi = 0,317), Asplenium sp. (Bi = 0,167), G. erecta (Bi = 0,096) y Rhynchospora sp. (Bi = 0,069). Respecto a los tallos, se encontraron fragmentos de nueve de las especies vegetales analizadas, donde sin embargo únicamente Asplenium sp. (Bi = 0,482), Anthurium sp. (Bi = 0,247) y Phytolacca sp. (Bi = 0,139) sobresalen por ser preferidos relativamente por éste roedor. Las Ericaceas son la mayor fuente de frutos, aún cuando se encontró una preferencia relativa superior hacia Macleania rupestris (Bi = 0,181) y C. nitida (Bi = 0,134) y en menor grado hacia Th. floribunda (Bi= 0,101). Sin embargo, en este caso, las diferencias en el índice de selección se pueden encontrar ligadas a los períodos de fructificación propios de cada especie. Adicionalmente, C. taczanowskii registró una elevada preferencia hacia los frutos de Phytolacca sp. (Bi = 0,457). En el caso de los rizomas, indudablemente los helechos son la mayor fuente disponible de este recurso alimentario, aunque Asplenium sp. (Bi = 0,685), Blechnum sp. (Bi= 0,159) y Cyathea sp. (Bi = 0,121) fueron identificados como especies preferidas en la dieta.
Esta selectividad de forrajeo, de acuerdo a la parte de la planta consumida, se observó igualmente al analizar el hecho de que los cuatro fragmentos de bosque presentaron diferencias en la composición florística con relación al recurso alimenticio evaluado. Es así como en los bosques (B1) y (B4) no se encuentra representada Anthurium sp., mientras que Th. floribunda y Phytolacca sp. están ausentes en (B2) y Cyathea sp. no se registró para el bosque (B4). Sin embargo, los fragmentos vegetales recuperados de la materia fecal colectada en los cuatro bosques no coinciden en todos los casos con la presencia o ausencia del recurso alimenticio (tabla 1), siendo extremo el hecho que en el bosque (B3) aunque está presente el recurso no se encontró materia fecal de la especie, es decir, C. taczanowskii aunque consume con mayor o menor preferencia las 16 especies de plantas evaluadas, se desplaza a través de los fragmentos de bosque seleccionando los recursos alimenticios disponibles de acuerdo a la presencia de frutos de Phytolacca sp. y de hojas, como es el caso de
Esta situación corrobora la hipótesis planteada por Osbahr (2003) en el sentido de que C. taczanowskii selecciona el recurso alimenticio de acuerdo a las necesidades calóricas y requerimientos nutricionales cuantitativos, lo cual afirma, por una lado, la selectividad de la especie y, por el otro, su baja dependencia sobre las abundancias de los recursos disponibles en el hábitat. Sin embargo, el consumo de hojas, tallos, frutos y rizoma de una misma planta o de varias plantas puede depender del estado fenológico vegetal de las mismas, por lo que se figura 2 podría estar subestimando o sobrestimando ciertas partes de una misma planta, dependiendo de su ciclo reproductivo (Anthony & Smith 1974). Este hecho se evidencia en el caso del consumo de hojas de Chusquea sp. con un porcentaje en la dieta del 10,39%, recurso de mayor preferencia por parte de C. taczanowskii (figura 2), por hallarse esta planta en su fase de floración durante el período de muestreo y, por lo tanto, con mayor número de rebrotes con hojas jóvenes de mejor palatabilidad, con la posibilidad de ser digerido.
Al analizar la posibilidad que la especie seleccione el alimento de acuerdo a la abundancia del mismo en el medio, se observa que los índices de selección no parecen estar ligados a plantas con mayores abundancias totales en el hábitat. Esta afirmación, se confirma al realizar el análisis de correlación de Pearson entre las dos variables, el cual indica que no existe una relación estadísticamente significativa entre las dos variables (r = -0,059 F(1,17) = 0,060 n = 19), indicando que la selección del recurso alimenticio es explicada únicamente en el 6% de los casos por la abundancia en el hábitat. Aunque la distribución espacial ligada a las condiciones ambientales y al grado de intervención del bosque favorece la formación de parches alimenticios con abundancias elevadas de determinado recurso (Osbahr, 2004), no necesariamente es el factor explícito para que la especie lo seleccione en mayor o menor grado.
Teniendo en cuenta los análisis fecales y las proporciones de los ítems alimentarios identificados mediante la técnica microhistológica, se encontró que, según el índice estandarizado de Hurlbert (1978), C. taczanowskii mostró una amplitud de nicho relativamente baja (BA´) = 0,247), lo cual indica que la especie discrimina entre los estados de recursos evaluados, tendiendo a ser selectiva (Krebs, 1999). Esta selectividad se encuentra ligada al consumo diferencial de los diferentes estados fenológicos de las plantas, discriminando en menor grado las hojas (BA´ = 0,235) y los frutos (BA´ = 0,136), mientras que es más especializado en el consumo de los tallos (BA´) = 0,099 y de los rizomas (BA´) = 0,064.
CONCLUSIONES
La relación no significativa entre el índice de selección y el recurso disponible corroboran que la especie no selecciona los recursos alimentarios de acuerdo a su abundancia en el hábitat.
De acuerdo a los índices de selección y de amplitud de nicho C. taczanowskii es una especie que utiliza de manera selectiva los estados fenológicos de las plantas, desplazándose grandes distancias entre los bosques muestreados en el Cerro Manjuí, en búsqueda de alimento.
Plantas como Th. floribunda, Phytolacca sp. y Asplenium sp. constituyen un recurso alimentario importante para la especie, debido a que son consumidas en diferentes estados fenológicos.
AGRADECIMIENTOS
Los autores agradecen a Guillermo Corredor por brindar la posibilidad de desarrollar la investigación en la Finca El Manjuí y a Mateo Hernández por su colaboración durante la colecta y la determinación del material vegetal.
BIBLIOGRAFÍA
1. AMAYA, J.A.; RODRÍGUEZ, A. 2004. Aspectos etológicos del tinajo de páramo (Agouti taczanowskii) en cautiverio. Trabajo de grado. Universidad Distrital Francisco José de Caldas, Facultad de Ciencias de Educación, Bogotá, 100p.
2. ANTHONY, R.G.; SMITH, N.S. 1974. Comparison of rumen and fecal analysis to describe deer diets. J. Wildlife Manamgement. 38(3):535-540
3. BAILLIE, J. 1996. Agouti taczanowskii. En: 2006 IUCN Red List of Threatened Species. Disponible desde Internet en: http://www.iucnredlist.org (con acceso 08/0307).
4. BARTOLOMÉ, J.; FRANCH, V.; GUTMAN; M.; SELIGMAN, V. 1995. Technical note: Physical factors that influence fecal analysis estimates of herbivore diets. J. Range Management. 48(3):267-270.
5. BATZLI, G.O.; COLE, F.R. 1979. Nutritional Ecology of Microtine Rodents: Digestibility of forage. J. Mammalogy. 60:740-750.
6. CASTRO, J.J. 1996. Estudio del Tinajo o Borugo (Agouti taczanowskii) (Agoutidae: Rodentia) en cautiverio. En: Seminario Internacional sobre Investigación, conservación y manejo de Agouti paca y Agouti taczanowskii comúnmente llamados Boruga, Guartinaja o Lapa. Santa Marta. Colombia. 40p.
7. COLE, F.R.; LOOPE, L.L.; MEDEIROS, A.C.; RAIKES, J.A.; WOOD, C.S. 1995. Conservation implications of introduced game birds in high-elevation hawaiian shrubland. Conserv. Biol. 9:306-313.
8. CORPORACIÓN AUTÓNOMA REGIONAL DE CUNDINAMARCA CAR. 2005. Formulación participativa del Plan Integral de Manejo del Distrito de Manejo Integrado de los Recursos Naturales Renovables del Sector Salto del Tequendama-Cerro Manjuí. Andean Geological Services LTDA. Bogotá DC. 1266p.
9. CORTÉS, A.; RAU, J.R.; MIRANDA, E.; JIMÉNEZ, J.E. 2002. Hábitos alimenticios de Lagidium viscacia y Abrocoma cinerea: roedores sintópicos en ambientes altoandinos del norte de Chile. Rev. Chilena de Historia Natural. 75:583-593.
10. DAJOZ, R. 2002. Tratado de Ecología. 2? edición. Ed. Mundiprensa, 510p.
11. DANIEL, W. 2004. Bioestadística: base para el análisis de las ciencias de la salud. 4? Edición. Limusa. México. 755p.
12. DELGADO, B.I.; O?ATE, O.L.; NIETO, J.A. 1991. Algunos aspectos bioecológicos de la guagua de páramo (Agouti taczanowskii) y perspectivas de su manejo en zoocriadero. Trabajo de grado. Universidad de la Salle, Facultad de Ciencias de la Educación, Departamento de Química y Biología, Bogota, 120p.
13. DRICKAMER, L. 1971. Experience and selection behavior in the food habits of Peromyscus: use of olfaction. Behaviour 61:267-287.
14. ECOTECNIA, 1997. Estudio de caracterización de cadenas suscesionales del hábitat para la Guagua (Agouti taczanowskii) en la zona amortiguadora del Parque Nacional Natural los Nevados. Informe técnico Corpocaldas, Manizales. 96p.
15. ETTER, A.; WYNGAARDEN, V. 2000. Patterns of Landscape Transformation in Colombia, with Emphasis in the Andean Region. AMBIO: A Journal of the Human Environment. 29(7):432-444.
16. GAVI?O, G.; JUÁREZ, C.; FIGUEROA, H.H. 1974. Técnicas biológicas selectas de laboratorio y de campo. Ed. Limusa. México. 251p.
17. GUERRERO, S.; BADII, M.; ZALAPA, S.; FLORES, A. 2002. Dieta y nicho de alimentación del coyote gris, mapache y jaguarundi en un bosque tropical caducifolio de la costa Sur del estado de Jalisco, México. Acta Zoológica Mexicana. 86:119-137.
18. HARTMAN, G. 1994. Long-Term Population Development of a Reintroduced Beaver (Castor biber) population in Sweden. Conserv. Biol. 8:713-717.
19. HOLECHEK, J.L.; VAVRA, M.; PIEPER, R.D. 1982. Botanical Composition Determination of Range Herbivore Diets: A review. J. Range Management 35: 309-315.
20. HURLBERT, S.H. 1978. The measurement of niche overlap and some relatives. Ecology. 59:67-77.
21. IUCN, 2007. 2007 IUCN Red List of Threatened Species. Disponible desde Internet en: http://www.iucnredlist.org (con acceso el 25/10/07).
22. JOHNSON, D.H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65-71.
23. KRONFELD, N.; DAYAN, T. 1998. A new method of determining diets of rodents. J. Mammalogy. 79:1198-1202.
24. KREBS, C.H. 1999. Ecological Methodology. 2nd Ed. Menlo Park. California. 620p.
25. LEE, G.J.; MACGREGOR, C.M. 2004. Comparison of a microhistological analysis of faeces and alkane concentrations of faeces to estimate the botanical composition of the diet of grazing sheep. Animal Production in Australia. 25:108-111.
26. LOU, S.; YURRITA, C. 2005. Análisis del nicho alimentario en la comunidad de murciélagos frugívoros de Yaxhá, Petén, Guatemala. Acta Zoológica Mexicana 21(1):83-94.
27. MANLY, B.F.J.; McDONALD, L.L.; THOMAS, D.L. 1993. Resource selection by animals: Statistical design and analysis for field studies. Chapman & Hall, London, 595p.
28. MESERVE, P.L. 1981. Trophic relationship among small mammals in a Chilean semiarid thorn scrub community. J. Mammalogy. 62: 304-314.
29. NÚ?EZ, C.; JORGENSON, J. P. 1996. Uso del hábitat natural y distribución de la población de ?borugo de páramo? Agouti taczanowskii, en relación con la alteración de los ecosistemas de alta monta?a, en el Parque Nacional Natural Chingaza. Trabajo de grado. Pontificia U. Javeriana. Fac. de Ciencias. Depto. Biología. Bogotá DC. 137p
30. OSBAHR, K. 1995. Distribución geográfica de tres especies de roedores histricomorfos en la región andina de Colombia. Informe técnico. Programa de Becas para la conservación. WCI, FES. GEA. 120p.
31. OSBAHR, K. 1996a. Disponibilidad de recursos alimenticios para el tinajo de páramo (Agouti taczanowskii) en el bosque nublado altoandino relictual en la región del alto del vino (Cundinamarca). Informe final. FONDO FEN. UDCA. Bogotá DC. 111p.
32. OSBAHR, K. 1996b. Evaluación preliminar de las causas de presión de origen antrópico sobre las poblaciones naturales de Agouti taczanowskii. En: Memorias Seminario Internacional sobre Investigación, Conservación y Manejo de Agouti paca y Agouti taczanowskii, comúnmente llamados Boruga, Guartinaja o Lapa. Santa Marta, Colombia. 40p.
33. OSBAHR, K. 1999. Identificación de plantas consumidas por Agouti taczanowskii y Dinomys branickii a partir de fragmentos vegetales recuperados en heces. Rev. U.D.C.A Act. & Div. Científica. 2:42-49.
34. OSBAHR, K. 2003. Patrones de distribución del recurso alimenticio disponible para Agouti taczanowskii (Rodentia, Agoutidae) en un fragmento de Bosque Andino Nublado. Rev. U.D.C.A Act. & Div. Científica. 6(1):38-48.
35. OSBAHR, K. 2004. Caracterización ambiental de un bosque andino nublado relictual y su relación con la densidad del recurso alimenticio disponible para Agouti taczanowskii. Rev. U.D.C.A Act. & Div. Científica. 7(2):73.84.
36. PARDO, A. 2004. Caracterización y uso de hábitat de Agouti taczanowskii en la Reserva Forestal Protectora Río Blanco (Cundinamarca). Trabajo de grado. Pontificia U. Javeriana, Fac. Ciencias. Depto. Biología. Bogotá, 77p.
37. PARRA, A.; OSBAHR, K. 2000. Contribución al conocimiento anatómico del aparato digestivo del tinajo de páramo (Agouti taczanowskii, Stolzman 1865). Rev. U.D.C.A Act. & Div. Científica. 3:21-27.
38. PARRA, A.; MARTÍNEZ. A.; JARAMILLO, R. 1970. Estudio general de suelos, municipios para fines agrícolas de las cuencas media y baja del río Bogotá y municipios aleda?os (Depto. Cundinamarca). Instituto Geográfico ?AGUSTIN CODAZZI?. Volumen VI. N°6. Bogotá, 550p.
39. PÉREZ-TORRES, J. 2002. Un índice para la evaluación del hábitat de Agouti taczanowskii(Rodentia: Agoutidae) en áreas de bosque andino nublado. Universitas Scientiarum. 7:51-60.
40. RANDOLPH, J.C.; CAMERON, G.N.; WRAZEN, J.A. 1991. Dietary choice of a generalist grassland herbivore, Sigmodon hispidus. J. Mammalogy. 72(2):300-313.
41. REICHMAN, O.J. 1975. Desert rodent diets. J. Mammalogy 56: 731-735.
42. SÁNCHEZ, F.A.; CADENA, A.; SÁNCHEZ, P. 1999. Abundancia, diversidad y uso de hábitat de grandes mamíferos em bosque andino y reforestado em Manizales (Caldas, Colombia). Trabajo de grado, U. N. Colombia, Fac. Ciencias, Depto. Biología, Bogotá. 98p.
43. SPARKS, D.R.; MALECHEK, J.C. 1968. Estimating percentage dry weight in diets using a microscopic technique. J. Range Management. 21:264-26.
44. STORR, G.M. 1961. Microscopic analysis of feces, a technique for ascertaining the diet of herbivorous mammals. J. Biol. Sci. 14:157-164.
45. TOBIN, M.E.; KOEHLER, A.E.; SUGIHARA, R.T. 1994. Seasonal patterns of fecundity and diet of roof rats in Hawaiian macademia orchards. Wildlife Res. 21:519-526.
Recibido: Agosto 12 de 2007; Aceptado: Noviembre 6 de 2007

Revista U.D.C.A Actualidad & Divulgación Científica por Universidad de Ciencias Aplicadas y Ambientales se distribuye bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.