CIENCIAS AGROPECUARIAS - Artículo Científico
EFECTO DE DIETAS MERÍDICAS EN LA TOXICIDAD DE LA PROTEÍNA CRISTALINA (Cry) DEL Bacillus thuringiensis SOBRE TRES PLAGAS DEL ALGODONERO (LEPIDOPTERA: NOCTUIDAE)
EFFECTS OF MERIDIC DIETS ON THE Bacillus thuringiensis CRYSTALINOUS PROTEINS (Cry) TOXICITY UPON THREE COTTON PESTS (LEPIDOPTERA: NOCTUIDAE)
Helber Arévalo1 Ingeborg Zenner de Polanía2 Laura Romero3
1Ingeniero Agrónomo. Programa jóvenes investigadores e innovadores ?Virginia Gutiérrez de Pineda? convenio Colciencias-U.D.C.A, Calle 222 No. 54-37, Bogotá D.C. Correo electrónico:harevalo@udca.edu.co
2I.A. Ph.D. Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A Correo electrónico:izenner@udca.edu.co
3Estudiante Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A Correo electrónico:lauraromeror@hotmail.com
Rev. U.D.C.A Act. & Div. Cient. 14(1): 39 - 48, 2011
RESUMEN
Desde la introducción de los cultivos genéticamente modificados (transgénicos) alrededor del mundo, se ha venido realizando un esfuerzo para monitorear la susceptibilidad de las diferentes especies que son el objetivo a controlar por parte de las toxinas Cry de estos cultivos. Sin embargo, difícilmente estos resultados pueden ser comparados de una región a otra, no sólo por la procedencia de las poblaciones insectiles, si no por el empleo de diversas metodologías; entre ellas encontramos el uso de dietas merídicas de componentes diferentes. El objetivo de este estudio, fue evaluar el efecto de tres dietas merídicas en la toxicidad de las proteínas cristalinas producidas por la bacteria B. thuringiensis sobre tres plagas del algodonero de El Espinal, Tolima. En todos los casos la menor susceptibilidad se presentó en la dieta ICRISAT, la cual ha exhibido los mejores resultados para la cría de estas plagas bajo nuestras condiciones. Los resultados encontrados no permiten inferir sobre un aumento o disminución de resistencia de la población de Heliothis virescens debido a que la cohorte evaluada fue recolectada sobre Desmodium sp.; para Spodoptera frugiperda, no se encontró dilución, ni adquisición de resistencia, mientras que en Helicoverpa zea se observa un leve aumento en la susceptibilidad. Los datos encontrados sugieren que se debe continuar con el monitoreo de la susceptibilidad de estas especies usando en lo posible la misma metodología para evitar una variación en los resultados y de igual manera emplear poblaciones procedentes de distintas geografías y plantas hospederas.
Palabras clave: Dietas merídicas, Toxinas Cry, Spodoptera frugiperda, Helicoverpa zea, Heliothis virescens.
SUMMARY
Since the introduction of genetically-modified (transgenic) crops around the world, an effort to monitor the susceptibility to Cry toxins on targeted insects has been implemented. However, these results are hardly comparable from region to region, not only due to the use in strains from different parts, but also due to methodological differences. Among these discrepancies is the use of meridic diets containing different ingredients. The goal of this study was to evaluate the effect of three meridic diets on the toxicity of crystal proteins from the bacterium B. thuringiensis on three cotton pests from El Espinal and Tolima. In all the tests, the lowest susceptibility was obtained with the ICRISAT diet, which in turn, it is superior for rearing these pests under our environmental conditions. The results also suggest that there is an increase or dilution of resistance in Heliothis virescens populations due to the fact that these insects were collected from Desmodium sp. Spodoptera frugiperda?s susceptibility was not affected, nor acquired resistance, while Helicoverpa zea had a slightly increase in susceptibility. These results suggest that Btsusceptibility should continue using the same methodology to decrease variability as well as tests to be conducted on populations from different areas and host plants.
Key words: Meridic diets, Cry toxins, Spodoptera frugiperda, Helicoverpa zea, Heliothis virescens.
INTRODUCCIÓN
En Colombia, al igual que en otros países, el uso de los cultivares que han sido genéticamente modificados (transgénicos) ha logrado disminuir la aplicación de insecticidas (Molina-Ochoa et al. 2010; Zenner de Polanía et al. 2008; Greenberg &Adamczyk, 2010); no obstante, la probabilidad que los noctuidos Helicoverpa zea (Boddie) y Heliothis virescens (Fabricius) adquieran resistencia, así como el bajo o nulo control de los cultivos transgénicos sobre Spodoptera frugiperda (J.E.Smith), aunado con la posibilidad de que otras plagas lepidóptera se puedan convertir en plagas primarias por la reducción en el número de aplicaciones de insecticidas y por la competencia interespecífica (Santos et al. 2009; Zenner de Polanía et al. 2008), hace que estos insectos sean objetivo de estudio en nuestro sector agrícola.
Además de la importancia mencionada, el aumento en la implementación del algodonero transgénico en Colombia, donde el área sembrada pasó de ser del 5% en el 2003 a más del 70% en la actualidad (Agro-Bio, 2009), hace necesario monitorear la respuesta de las diferentes poblaciones a las toxinas Cry presentes en el algodonero Bt, con el fin de conocer la evolución de la resistencia de los insectos sujetos a control por esta tecnología. Conociendo esta respuesta, se puede prevenir a los agricultores para que estén preparados y puedan tomar a tiempo las medidas pertinentes (Zenner de Polanía et al. 2008).
Para monitorear la resistencia de estas plagas a las toxinas Cry, se emplea la técnica del bioensayo, en la cual, se exponen larvas neonatas, bajo condiciones de laboratorio, a la toxinas Cry del Bt, empleando un alimento artificial; sin embargo, los bioensayos elaborados, hasta el momento, presentan algunas diferencias, que afectan los datos obtenidos. Las variaciones más comunes giran en torno a cuatro aspectos. 1) La obtención de las poblaciones insectiles: los investigadores parten de poblaciones recolectadas de diferentes plantas hospederas en el campo o a partir de poblaciones criadas bajo condiciones de laboratorio. En este aspecto, se ha comprobado que la respuesta es diferente, cuando las poblaciones provienen de distintas partes geográficas (Ali et al. 2006) o cuando las poblaciones provienen de diferentes plantas hospederas (Zenner de Polanía et al. 2009). También, cuando las cohortes son obtenidas de laboratorios, lo que ha permitido la elaboración de la técnica de isofamilias en la que se ha estudiado la condición genética de la resistencia (Blanco et al. 2010). 2) La forma de obtención de las diferentes toxinas Cry: debido a que se puede partir de tres maneras diferentes purificando la toxina: a partir de plantas transgénicas; a partir de formulaciones comerciales, o purificando la toxina, a partir de cepas clonadas de E. coli recombinante (Akhurst et al. 2003). 3) La disposición de las toxinas en la dieta merídica: dado a que en algunos casos se imposibilita incorporar altas concentraciones de la toxina en la dieta, lo que ha obligado a los investigadores a adicionar la toxina solamente en la superficie de la misma (Akhurst et al. 2003). 4) Al empleo de diferentes dietas artificiales para la cría de insectos, debido a que la adquisición de dietas específicas o de algunos ingredientes se dificulta, ya sea porque no se encuentran en el mercado donde se realiza el estudio o son demasiado costosos. Sobre esta variable, Blanco et al. (2009) han comprobado cómo se puede encontrar una respuesta diferencial, debido a la cantidad de alimento y, con ello, de toxina consumida y que algunos ingredientes puedan inferir en la activación de las protoxinas Cry.
Uno de los puntos más críticos en el empleo de las dietas merídicas es, por lo tanto, su costo, por lo cual, muchos investigadores se han dedicado al desarrollo de dietas de bajo costo, buscando ingredientes más baratos, eliminado ingredientes, aparentemente no esenciales y evaluando sustitutos potenciales para el agar (Morimoto et al. 2004; Cappellozza et al. 2005). Todas estas variables impiden la comparación de los resultados entre regiones y hacen necesario que se continúe con la evaluación del efecto que tiene cada una de estas variaciones, en la obtención de resultados.
El empleo de dietas con diferentes componentes no solamente puede tener un efecto sobre los resultados obtenidos en los bioensayos para monitorear resistencia o las crías masivas de insectos, sino también en la producción de, por ejemplo, hongos entomopatógenos o virus usados en control biológico. Elvira et al. (2010) evaluaron el efecto de la dieta consumida por Spodoptera exigua sobre la producción del virus de la nucleopolihedrosis y no hallaron diferencias entre una dieta de bajo costo y otra comúnmente usada.
Por esta razón, el objetivo de este estudio fue determinar el efecto de tres dietas merídicas en la toxicidad de las toxinas Cry y evaluar la respuesta de diferentes poblaciones a las toxinas.
MATERIALES Y MÉTODOS
La investigación, se efectuó en el Laboratorio de Biotecnología Agrícola, Facultad de Ingeniería Agronómica, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A. Las condiciones ambientales fueron 25,6±2,3°C y 70±5% de humedad relativa.
Tres poblaciones de Spodoptera frugiperda en estado larval fueron obtenidas del municipio El Espinal, Tolima; una, del laboratorio de Biotecnología Agrícola de la Universidad de Ciencias Aplicas y Ambientales U.D.C.A, originalmente recolectada sobre maíz convencional, la cual, había sido criada por cinco generaciones; la segunda obtenida también de maíz convencional y, la tercera, se recolectó sobre algodonero transgénico. Tanto en la segunda como en la tercera población, se empleó la primera generación obtenida en laboratorio. La población de H. virescens fue extraída del laboratorio de Biotecnología Agrícola de la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, recolectada sobre pega-pega (Desmodium sp.). La colonia de H. zea, se inició a partir de larvas de los dos últimos instares obtenidas de mazorcas de maíz atacadas. Las larvas de las diferentes colonias fueron criadas una generación antes de iniciar el estudio sobre la dieta ICRISAT diet 3, a base de garbanzo, preparada de acuerdo a la fórmula proporcionada por el doctor H. Sharma del Instituto Internacional de Investigación en Cultivos para las Zonas Tropicales Semiáridas (ICRISAT), en Patancheru, Andhra Pradesh, India, la cual, presentó los mejores resultados biológicos para S. frugiperda y H. virescens bajo nuestras condiciones de cría (Arévalo & Zenner de Polanía, 2009; 2010).
Larvas neonatas, se obtuvieron del apareamiento de diez parejas puestas en una cámara de cría de vidrio de 3,7L. Los adultos fueron alimentados con la solución azucarada, sugerida por Greene et al. (1976), consistente en 1g de miel, 0,1g de ácido ascórbico, 0,1g de methyl parahydroxybenzoato y 6g de azúcar, diluidos en 100mL de agua, que se brindó humedeciendo una mota de algodón diariamente. Las cámaras de cría fueron tapadas con tela tole y, para la oviposición, se dispusieron tiras de papel toalla desde la parte superior, hasta el fondo. Los huevos fueron transferidos, a diario, a un vaso limpio, removiéndolos de la tela o del papel toalla, con una solución al 0,26% de hipoclorito de sodio (Akhurst et al. 2003).
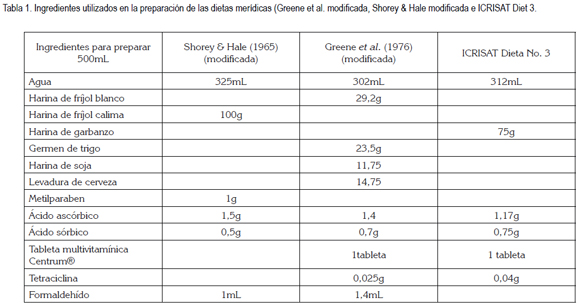
Las dietas empleadas en este estudio fueron: 1) ICRISAT Diet 3; 2) Greene et al. (1976) modificada por Parra (2001) (García et al. 2006); 3) Shorey &Hale (1965), modificada por Bowling (1967) y preparada según Villacorta &Cobo (1978) (Tabla 1).
El Cry1Ab empleado en los bioensayos fue multiplicado, a partir de la cepa ECE54, suministrada por el Bacillus Genetic Stock Center. La cepa está compuesta por la proteína Cry1Ab clonada dentro de células de Escherichia coli cepa E. coli JM103, con el vector de expresión pKK223-3 (Ge et al. 1989).
Para la multiplicación de la cepa, se siguió la metodología de Akhurst et al. (2003), con algunas modificaciones, para aumentar la concentración de la toxina, descritas a continuación. La cepa de E. coli, que expresa el gene del Cry1Ab, fue cultivado en 10mL de caldo Luria-Bertani con 100µg/mL de ampicilina, por 24 horas, a 37°C; posteriormente, se aumentó la producción, llevando esta mezcla a un litro del Luria-Bertani con 100µg/mL de ampicilina, por tres días, a 37°C, en agitación permanente; luego de completado este tiempo, la muestra fue centrifugada a 4000rpm, por 20 minutos, a temperatura ambiente. El supernadante fue decantado y el pellet resuspendido en agua destilada. La suspensión fue sonicada para dispersar completamente los cristales, en dos tratamientos de 5min, con un periodo intermedio de 5min, a 4°C.
El Cry1Ac, se obtuvo del insecticida MVP II® (Cry1Ac encapsulado producido por Pseudomonas sp., proporcionado por Dow CropSciences). Para establecer la concentración de las proteínas, se utilizó el método de Bradford, con ayuda de un espectrofotómetro Spectronic 601 digital, a una longitud de onda de 565nm (Scopes, 1987). Antes de incorporar las respectivas proteínas en las dietas, se corroboró la presencia de las toxinas con el Kit para toxinas Cr1Ac y Cry1Ab QualiPlate de EnvirologixTM (Envirologix, 2007).
La susceptibilidad de las larvas neonatas, se evaluó incorporando diferentes concentraciones del Cry1Ac y Cry1Ab a las dietas merídicas, seis dosis de Cry1Ac entre 0,01 y 100µg/mL de dieta para H. virescens y H. zea. Las dosis, se establecieron de acuerdo a las CL50 de 3,52-3,81µg/ mL, para H. virescens y de 3,42-6,12 µg/mL, para H. zea, encontradas por Zenner de Polanía et al. (2008; 2010). En el caso de S. frugiperda (cogollero del maíz), se aplicaron siete dosis de Cry1Ac, que variaron entre 0,01 y 1000µg/ mL de dieta, escogidas de acuerdo a los reportes de los autores mencionados, de una CL50 máxima de 1178µg/mL. Se emplearon ocho dosis de Cr1Ab entre 0 y 15µg/mL, para evaluar la respuesta de S. frugiperda a esta toxina, que incluye, la CL50 (5,64-8,35µg/mL), hallada por Zenner de Polanía et al. (2009). En todos los bioensayos, se emplearon 30 larvas de forma individual por dosis. La mortalidad, se estimó siete días después y los datos se sometieron a análisis Probit (SAS Institute Inc.), para determinar las concentraciones letales. Con valores obtenidos desde el análisis Probit, se realizó un análisis de varianza, con el fin de determinar si se presentaban diferencias entre las concentraciones letales obtenidas en cada una de las dietas. Cuando se revelaron diferencias, las medias fueron separadas por el procedimiento de Duncan (Blanco et al. 2009).
RESULTADOS Y DISCUSIÓN
En la tabla 2, se puede observar cómo la colonia de S. frugiperda, criada por cinco generaciones bajo condiciones de laboratorio, mantiene una relativamente baja susceptibilidad a la toxina Cry1Ac en las dietas Greene y Shorey &Hale, mientras que en la dieta ICRISAT, se denota una menor susceptibilidad, que fue, incluso, menor a la encontrada en el 2005 y 2006,en poblaciones procedentes de los Departamentos de Tolima y de Cundinamarca (Zenner de Polanía et al. 2008), utilizando esta misma dieta.
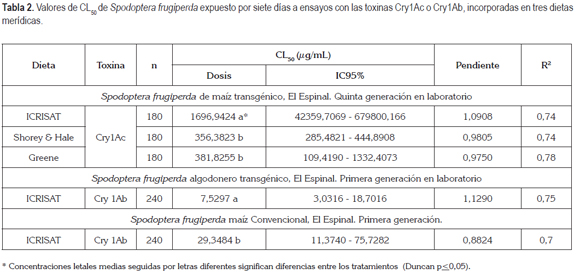
Sin importar las variaciones en la realización de los bioensayos, el cogollero ha mostrado una baja sensibilidad a la toxina Cry1Ac del Bt., ya sea bajo condiciones de laboratorio como de campo (Blanco et al. 2010), lo que sugiere que esta especie es resistente a la toxina Cry1Ac. Esta resistencia, se pudo haber desarrollado por la exposición del cogollero de forma natural al B. thuringiensis, generando variaciones, incluso en su genética (Monnerat et al. 2006). Una muestra de la variabilidad genética, se ha observado en otras especies al encontrarse receptores alterados en el intestino medio (Storer et al. 2010), como los reportados por Jurat-Fuentes &Adang (2004), indicando una correlación de la alteración de la glycolisación en el sitio de la inserción de las proteínas y la resistencia de una colonia de H. virescens a la toxina Cry1Ac.
En la misma tabla 2, se puede observar la respuesta al Cry1Ab de las poblaciones de S. frugiperda procedentes de maíz convencional y de algodonero transgénico. Los resultados obtenidos complementan el estudio de susceptibilidad de esta especie, procedente del Tolima, a la toxina Cry1Ab, desde el 2006, encontrándose que la población proveniente del algodón transgénico mantiene su susceptibilidad a la toxina, estando muy cercano a los valores de la línea base de 5,64µg/mL. En contraste, se puede observar cómo la población procedente de maíz convencional presenta una menor susceptibilidad y se observa un incremento con relación a la línea base, ya mencionada (Zenner de Polanía et al. 2009), sugiriendo que se debe mantener el monitoreo de esta especie y, en lo posible, emplear en el monitoreo poblaciones procedentes de diferentes cultivos y posiciones geográficas.
Los resultados hallados para Cry1Ac y Cry1Ab corroboran la incidencia de las poblaciones de Spodoptera spp. en los cultivos transgénicos, sobre todo en aquellos órganos donde la concentración de la toxina es menor (Santos et al. 2009; Zenner de Polanía et al. 2008). A pesar de este hecho es necesario buscar una medida para prolongar la vida útil de los cultivares Bt, debido a que éstos han cumplido con algunos de sus objetivos, como la disminución de las pérdidas y de la disminución del número de aplicaciones de insecticidas, incluso, en cultivos con presencia de poblaciones de Spodoptera frugiperda. Aunque en Colombia no se tienen estimativos de las poblaciones de S. frugiperda es posible que estas hayan disminuido, desde la introducción de los cultivares Bt, no por causar la mortalidad de su estado inmaduro sino por afectar su desarrollo.
En el caso de H. virescens, el gene Cry1Ac, incorporado en el algodonero, continua ofreciendo un control efectivo de esta plaga en Colombia e, incluso, se puede evidenciar en la figura 1, cómo la dosis necesaria para controlar la mitad de la población es menor a la registrada en 2005 y 2006 por Zenner de Polanía et al. (2008). A pesar de que la procedencia de la población evaluada es similar a la empleada en 2005 y 2006, no se pude inferir sobre una dilución en la resistencia, debido a que los individuos fueron recolectados sobre la arvense pega-pega y no sobre algodonero transgénico o convencional.

A pesar de los resultados encontrados es vital no bajar la guardia y continuar con el monitoreo de la susceptibilidad de esta plaga a la toxinas Cry del Bt. Para realizar esta labor, se sugiere emplear poblaciones de los lotes refugios, de los lotes con algodonero transgénico y del hospedero alterno Desmodium sp. La importancia del monitoreo radica en la fluctuación poblacional que esta especie ha mostrado, a través del tiempo, presentándose en bajas poblaciones, luego de emplear una nueva herramienta de control y presentarse una explosión poblacional, después de un tiempo, generando, de esta manera, grandes pérdidas económicas, como fue el caso de la adopción de los piretroides (Zenner de Polanía et al. 2008).
Al igual que Blanco et al. (2009), se encontraron diferencias significativas entre las dietas empleadas, observándose una menor susceptibilidad cuando la toxina fue incorporada en la dieta ICRISAT; no obstante, estos autores anotan que la respuesta puede ser directamente proporcional a la cantidad consumida, pero, en este estudio, no se halló la misma respuesta, debido a que precisamente la dieta ICRISAT es una de las más consumidas por esta especie.
La respuesta de H. zea (el gusano de la mazorca), durante dos generaciones a la toxina Cry1Ac, en tres dietas merídicas, se consigna en la figura 2. Los resultados presentan una diferencia significativa en la primera generación entre las dietas empleadas. Las diferencias obtenidas entre las dietas, no indican una tendencia, lo que dificulta el descarte o la adopción de una dieta; sin embargo, a pesar de ir en contra de lo esperado, se muestra una disminución en la susceptibilidad en la segunda generación, criada bajo condiciones de laboratorio, que puede estar relacionada con la adaptación y la nutrición del insecto a las dietas merídicas. Esto, se podría interpretar que la dieta, al suministrarle los mejores nutrientes al insecto, ayuda a que éste tenga un mejor desarrollo y sea menos susceptible a la toxina.

Las CL50 encontradas en ambas generaciones de H. zea presentaron una mayor susceptibilidad a la reportada por Zenner de Polanía et al. (2010), que varió entre 3,45 y 6,12µg/mL, permitiendo inferir, que se ha presentado un leve aumento de la susceptibilidad de esta especie al Cry1Ac.
Al igual que en las demás especies, la menor susceptibilidad de H. zea se presentó en la dieta ICRISAT, lo que corroboraría el hecho que, además de la cantidad de proteína y dieta consumida, la calidad putativa nutricional de la dieta afecta, de forma inversa, la susceptibilidad del insecto, es decir, a una mayor calidad de alimento la susceptibilidad del insecto a la toxina será menor, aunque es importante tener en cuenta cómo algunos ingredientes de las dietas pueden estar alterando la expresión de algunas enzimas digestivas (Blanco et al. 2010). Partiendo de esta premisa podríamos concluir que la dieta ideal para poder evaluar la susceptibilidad de la especie a las diferentes toxinas Cry, es aquella en la que el consumo y la presencia de nutrientes sean similares a la planta hospedera y, de la misma forma, que ninguno de los ingredientes altere la activación de la protoxina y la unión de la proteína insecticida con los receptores.
Las susceptibilidades halladas para S. frugiperda al Cry1Ab indican que los cultivos transgénicos, empleados actualmente, no le causan una mortalidad significativa, sobre todo en algunos órganos del maíz, donde la concentración es menor a 8µg/mL (Abel &Adamczyk, 2004; Mejía &Zenner de Polanía (datos sin publicar). Este hecho, se ha corroborado en campo, donde las larvas del cogollero se encuentran alimentándose, especialmente, en estas estructuras. En apariencia, los transgénicos están cumpliendo con parte de su propósito al disminuir el número de aplicaciones de insecticidas, al reducir costos y al reducir el daño; esto, ya que las concentraciones del Cry presentes en los tejidos del huésped afectan el desarrollo del insecto, provocando que se forme un menor número de generaciones por ciclo del cultivo y, en el caso del maíz, que el porcentaje de daño provocado en la mazorca sea menor, tal como lo describe Buntin et al. (2004).
Los datos obtenidos en este estudio permiten percatar cómo las dietas presentan una incidencia en la obtención de los resultados en las tres especies de insectos evaluadas, que no está determinado solamente por la cantidad de alimento consumido, sino por la calidad de éste y los efectos de algunos ingredientes en la activación de las protoxinas del Bt. También fue posible determinar cómo la procedencia del insecto afecta su susceptibilidad, lo cual, se podría atribuir a alteraciones genéticas por la exposición del insecto a diferentes entornos.
Además, se concluye que la susceptibilidad de H. zea y S. frugiperda no ha presentado una variación de la CL50 con respecto a la línea base, mientras que la susceptibilidad de H. virescens no puede ser comparada, debido a que la procedencia de la población evaluada es diferente a la población, con la cual, se generó la línea base.
Finalmente, se considera que para fines del monitoreo de la susceptibilidad a los Crys de las especies colombianas de noctuidos evaluadas, se debe aplicar la metodología establecida que emplea la dieta ICRISAT. Igualmente, al determinar las líneas base para otros Crys, incorporados al algodonero o al maíz, se debe respetar la metodología ya comprobada como eficiente.
AGRADECIMIENTOS
Al grupo de investigación ?Fitosanidad?, de la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A y a COLCIENCIAS, por la cofinanciación del presente trabajo. Conflictos de interés: El artículo fue preparado y revisado con la participación de todos los autores, quienes declaramos que no existe ningún conflicto de intereses que ponga en riesgo la validez de los resultados presentados.
BIBLIOGRAFÍA
1. ABEL, C.A.; ADAMCZYK, Jr., J.J. 2004. Relative concentration of Cr1A in Maize leaves and cotton bolls with diverse chlorophyll content and corresponding larval development of fall armyworm (Lepidoptera: Noctuidae) and southwestern corn borer (Lepidoptera: Crambidae) on maize whorl leaf profiles. J. Econ. Entomol. 97(5):1737-1744.
2. AGRO-BIO. 2009. Estadísticas de cultivos GM 2009. Disponible desde Internet en http://www.agrobio.org/ index.php?option=com_content&task=view&id=73 50&Itemid=15 (con acceso el 6/01/11).
3. AKHURST, R.J.; JAMES, W.; BIRD, L.J.; BEARD, C. 2003. Resistance to the Cry1Ac δ-endotoxin of Bacillus thuringiensis in the cotton bollworm Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 96(4): 1290-1299.
4. ALI, M.I.; LUTRELL, R.G.; YOUNG, III, S.Y. 2006. Susceptibilities of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae) populations to Cry1Ac insecticidal protein. J. Econ. Entomol. 99(1):164-175.
5. ARÉVALO-MALDONADO, H.A.; ZENNER DE POLANÍA, I. 2009. Evaluación de dietas merídicas para la cría en laboratorio de Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). Rev. U.D.C.A Actual. &Divul. Cient. 12(1):79-90. Disponible desde Internet en: http://www.scielo.org.co/pdf/rudca/v12n1/v12n1a09. pdf (con acceso el 10/11/10).
6. ARÉVALO-MALDONADO, H.A.; ZENNER DE POLANÍA, I. 2010. Evaluation of meridic diets suitable for efficient rearing of Heliothis virescens (F.) (Lepidoptera: Noctuidae). Rev. U.D.C.A Actual. &Divul. Cient. 13(2):163-173.
7. BLANCO, C.A.; GOULD, F.; VEGA-AQUINO,P.; JURATFUENTES, J.L.; PERERA, O.P.; ABEL, C.A. 2009. Response of Heliothis virescens (Lepidoptera: Noctuidae) strains to Bacillus thuringiensis Cry1Ac incorporated into different insect artificial diets. J. Econ. Entomol. 102(4):1599-1606.
8. BLANCO, C.A.; PORTILLA, M.; JURAT-FUENTES, J.L.; SÁNCHEZ, J.F.; VITERI, D.; VEGA-AQUINO, P.; TERÁN-VARGAS, A.P.; AZUARA-DOMINGUEZ, A.; LOPEZ JR., J.D.; ARIAS, R.; ZHU, Y.; LUGOBARRERA, D.; JACKSON, R. 2010. Susceptibility of isofamilies of Spodoptera frugiperda (Lepidoptera: Noctuidae) to Cry1Ac and Cry1Fa proteins of Bacillus thuringiensis. Southwestern Entomologist. 35(3):409-415.
9. BOWLING, C.C. 1967. Rearing of two lepidopterous pests of rice on a common artificial diet. Ann. Ent. Soc. Am. 60(6):1215-1216.
10. BUNTIN, D.G; ALL, J.N.; LEE, D.R.; WILSON, D.M. 2004. Plant-incorporated Bacillus thuringiensis resistance for control of fall armyworm and corn earworm (Lepidoptera: Noctuidae) in corn. J. Econ. Entomol. 97(5):1603-1611.
11. CAPPELLOZZA, L.; CAPPELLOZZA, S.; SAVIANE, A. 2005. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 40:405-412.
12. ELVIRA, S.; GORRÍA, N.; MUÑOZ, D.; WILLIAMS, T.; CABALLERO, P. 2010. A simplified low-cost diet for rearing Spodoptera exigua (Lepidoptera: Noctuidae) and its effect on S. exigua nucleopolyhedrovirus production. J. Econ. Entomol. 103(1):17-24.
13. ENVIROLOGIX. 2007. QualiplateTMkit for Cry 1Ab/Cry1Ac. Disponible desde Internet en: http://www.envirologix. com/library/ap003crbsinsert.pdf (con acceso el 15/11/10).
14.GARCÍA, M.S.; BUSATO, G.R.; GIOLO, F.P.; MANZONI, C; BERNARDI, O.C.; ZART, M.; NUNES, A.M. 2006. Tabela de vida de fertilidade de Helicoverpa zea (Boddie, 1850) (Lepidoptera: Noctuidae) em duas dietas artificiais. Rev. Bras. Agrociencia (Pelotas). 12(1):51-55. Disponible desde Internet en http:// www.ufpel.tche.br/faem/agrociencia/v12n1/artigo09. pdf (con acceso el 07/09/07).
15. GE, A.Z.; SHIVAROVA, I.; DEAN, H.D. 1989. Location of the Bombyx mori specificity domain on a Bacillus thuringiensis δ-endotoxin protein. Proc. Nal. Acad. Sci. 83(11):4037-4041.
16. GREENBERG, S.M.; ADAMCZYK, J.J. 2010. Effectiveness of transgenic Bt cottons against Noctuidae in the Lower Grande Valley of Texas. Southwestern Entomologist. 35(4):539-549.
17. GREENE, G.L.; LEPLA, N.C.; DICKERSON, W.A. 1976. Velvetbean caterpillar: a rearing procedure and artificial medium. J. Econ. Entom. Lanham. 69(4):488-497.
18.JURAT-FUENTES, J.; ADANG, M.J. 2004. Characterization of a Cry1Ac-receptor alkaline phosphatase in susceptible and resistant Heliothis virescens larvae. Europe Journal of Biochemestry. 217(15):3127-3135. Disponible desde Internet en http://onlinelibrary.wiley.com/doi/10.1111/j.1432- 1033.2004.04238.x/pdf (con acceso 1/04/10).
19. PMORIMOTO, M.; MATSUDA, K.; OHTA, Y. 2004. Evaluation of calcium-alginate gel as an artificial diet medium for bioassays on common cutworms. J. Agric. Food Chem. 52:4737-4739.
20. MEJÍA C., R.A.; ZENNER DE POLANÍA, I. 2011. Expresión de la toxina Cry1ab en maíz transgénico Yielgard® en los Llanos Orientales de Colombia (en evaluación).
21.MOLINA-OCHOA, J; HUTCHISONS, W.D.; BLANCO A., C. 2010. Estado actual de Helicoverpa zea y Heliothis virescens dentro de un panorama cambiante en el sur de los Estados Unidos de Norte América y México. Southwestern Entomologist. 35(3):347-354.
22. MONNERAT, R.; MARTINS, E.; QUEIROZ, P.; ORDÚZ, S.; JARAMILLO, G.; BENINTENDE, G.; COZZI, J.; REAL, M.D.; MARTINEZ-RAMIREZ, A.; RAUSELL, C.; CERÓN, J.; IBARRA, J.E.; RINCON-CASTRO, M.C.; ESPINOZA, A.M.; MEZA-BASSO, L.; CABRERA, L.; SÁNCHEZ, J.; SOBERON, M.; BRAVO, A. 2006. Genetic variability of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) populations from Latin America is associated with variations in susceptibility to Bacillus thuringiensis Cry toxins. Appl. Environ. Microbiol. 72: 7029-7035. Disponible desde Internet en: http://aem.asm.org/cgi/reprint/72/11/7029 (con acceso el 14/01/11).
23. ANTOS-AMAYA, O.; DELGADO-RESTREPO, O.; ARGÜELLES, J.; AGUILERA-GARRAMUÑO, E. 2009. Evaluación del comportamiento del complejo Spodoptera con la introducción de algodón transgénico al Tolima, Colombia. Revista Corpoica ? Ciencia y Tecnología Agropecuaria 10(1):24-32. Disponible desde Internet en http://www.corpoica.org.co/SitioWeb/Archivos/Revista/ 3.EvaluacindelcomportamientodelcomplejoSpodoptera. pdf (con acceso el 6/01/11).
24. SCOPES, R.K. 1987. Protein purification principles and practice, 2nd ed. Polyp.Springer. New York. p.280-282.
25. SHOREY, H.H.; HALE, R.L. 1965. Mass-Rearing of the larvae of nine Noctuid species on simple artificial medium. J. Econ. Entomol. 58(3):522-524.
26. STORER, N.P.; BABCOCK, J.M.; SCHLENZ, M.; MEADE, T.; THOMPSON, G.D.; BING, J.W.; HUCKABA, R.M. 2010. Discovery and characterization of field resistance to Bt Maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 103(4): 1031-1038.
27. VILLACORTA, A.; COBO DE MARTÍNEZ, L.S. 1978. Efecto del modo de preparación de la dieta sobre el crecimiento y desarrollo de Spodoptera frugiperda. ICA (Colombia). p.96-103.
28. ZENNER DE POLANÍA, I.; ÁLVAREZ-RODRÍGUEZ, J.A.; ARÉVALO-MALDONADO, H.A.; MEJÍA-CRUZ, R.; BAYONA-ROJAS, M. 2008. Susceptibilidad de cuatro noctuidos plaga (Lepidoptera) al gene Cry1Ac del Bacillus thuringiensis incorporado al algodonero. Rev. Colom. Entomol. 34(1):41-50. Disponible desde Internet en http://www.scielo.org.co/pdf/rcen/v34n1/ v34n1a05.pdf (con acceso el 6/01/11).
29. ZENNER DE POLANÍA, I.; ARÉVALO-MALDONADO, H.A.; MEJÍA-CRUZ, R.; DÍAZ-SÁNCHEZ, J. 2009. Spodoptera frugiperda: respuesta de distintas poblaciones a la toxina Cry1Ab. Rev. Colom. Entomol. 31(1):34-41. Disponible desde Internet en: http://www.scielo.org.co/scielo.php?script=sci_ arttext&pid=S0120-04882009000100007&lng=en& nrm=iso (con acceso el 13/10/09).
30. ZENNER DE POLANÍA, I.; ÁLVAREZ R., J.A.; ARÉVALO M., H.A. 2010. Respuestas de Helicoverpa zea (Lepidoptera: Noctuidae) a la toxina Cry1Ac del Bacillus thuringiensis. Southwestern Entomologist. 35(1):85-98.
Recibido: Enero 21 de 2011; Aceptado: Abril 13 de 2011

Revista U.D.C.A Actualidad & Divulgación Científica por Universidad de Ciencias Aplicadas y Ambientales se distribuye bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.