
CIENCIAS AGROPECUARIAS - Artículo Científico
COMPONENTES DE VARIANCIA EN VARIABLES DE CRECIMIENTO Y FOTOSÍNTESIS EN FRÍJOL COMÚN (Phaseolus vulgaris L.)
COMPONENTS OF VARIANCE IN GROWTH TRAITS AND PHOTOSYNTHESIS IN COMMON BEAN (Phaseolus vulgaris L.)
Gustavo A. Ligarreto M.1
1I.A. Ph.D., Profesor Asociado. Facultad de Agronomía. Universidad Nacional de Colombia. Bogotá. e-Mail: galigarretom@unal.edu.co
Rev. U.D.C.A Act. & Div. Cient. 16(1): 87 - 96, 2013
RESUMEN
Las colecciones de fríjol se han estudiado por características morfológicas y de producción, pero hace falta el conocimiento de los caracteres relacionados con el crecimiento de la planta y la fotosíntesis; la aplicación de esta investigación permite apoyar a los programas de mejora genética de cultivares de alto valor para la seguridad alimentaria. En una colección de fríjol común, compuesta por 29 materiales de procedencias Argentina, Bolivia, Colombia, México y Perú existente en el banco de germoplasma de la Nación colombiana que administra la Corporación Colombiana de Investigación Agropecuaria (Corpoica), se midieron 34 variables cuantitativas relacionadas con el crecimiento de la planta y la fotosíntesis, con el objetivo de seleccionar las variables de mayor aporte genético y poco efecto de ambiente. Los análisis para las variables cuantitativas de índole fisiológica revelaron alto efecto del ambiente sobre su expresión, excepto sobre los componentes de crecimiento razón área foliar (RAF), razón peso foliar (RPF) y área foliar específica (AFE), que presentaron mayor expresión genética, indicando que son caracteres que se pueden usar para describir las accesiones de la colección y como variables, con uso potencial, para la selección de genotipos, en programas de mejoramiento genético.
Palabras clave: Crecimiento y desarrollo, evaluación, germo- plasma, leguminosa comestible.
SUMMARY
Bean collections have been documented by morphological and production traits, but the knowledge related with growth and photosynthesis are deficient, allowing the application of this research to support breeding programs of high value cultivars for food security. In a common bean population of 29 accessions of the Colombian germplasm bank administrated by the "Corporación Colombiana de Investigación agropecuaria (Corpoica)" with origins in Argentina, Bolivia, Colombia, Mexico and Peru, 34 variables were measured with several quantitative characteristics related to plant growth and photosynthesis, in order to identify the variables with higher genetic improvement and lower environmental effects. The analysis of the 34 quantitative variables of physiologic nature revealed the high effect of the environment on their expression, except on the leaf area ratio (LAR), the leaf weight ratio (LWR) and the specific leaf area (SLA), which are more genetic expression, showing them as possible characters to be used at accessions description and as potential variables for the genotype selection in breeding programs.
Key words: Growth and development, evaluation, germplasm, food legume.
INTRODUCCIÓN
La descripción y la documentación de las colecciones de germoplasma que se relacionan en los datos de pasaporte constituyen el valor agregado de las accesiones, pues dejan reconocer su uso potencial, al igual que permite a los programas de recursos genéticos vegetales a conocer la variabilidad genética que conservan y sirven para que los programas de mejoramiento genético disminuyan el número de genotipos y el tiempo invertido en premejoramiento (Borém, 1998).
Durante las últimas décadas, se han aplicado múltiples estrategias tendientes a aumentar la producción de alimentos; sin embargo, las ganancias han sido evaluadas como simples métodos de incremento de los componentes del rendimiento, como peso del grano, el peso de la vaina, mazorcas, espigas y panojas, entre otras estructuras reproductivas y del número de granos por estructura, pero la producción es el resultado del componente genético y del ambiente, considerando el cultivar y el efecto de los factores bióticos y abióticos sobre su expresión. Por tanto, en cualquier intención de mejora genética, es importante considerar la posibilidad de manipular los genes involucrados en el rendimiento, en las resistencias a patógenos y en el estrés abiótico, en la alta calidad nutricional y en la eficiencia fisiológica, a través del uso de los recursos genéticos con genes de interés (Bressan & Pereira, 2003).
El fríjol presenta contenido de proteína, que varía del 20 al 28% y tiene aminoácidos esenciales, como metionina, lisina y triptófano, además de calcio y hierro; este alto valor nutricional ha hecho que la especie sea considerada la principal fuente de proteína para poblaciones de escasos recursos, especialmente, para países en desarrollo, donde el consumo per capita puede alcanzar los 50kg en algunas regiones (Broughton et al. 2003). Para el 2009, a nivel global, se cosechó un área de 25.563.866ha, con una producción de 20.698.984t, en el que participaron 121 países (FAO, 2011).
La producción de esta leguminosa podría estar seriamente afectada, puesto que el fríjol se considera como uno de los cultivos más vulnerables al cambio climático. Según el programa de investigación enfocado en cambio climático, agricultura y seguridad alimentaria (CCAAF), que hace parte del Grupo Consultivo para Investigación Agrícola Internacional (CGIAR), la generación de variedades que respondan a las nuevas y variables condiciones ambientales, se destaca como una de las soluciones para enfrentar y mitigar el efecto del cambio climático en la agricultura (Jarvis et al. 2011).
La descripción de los estados de desarrollo de las plantas, el estudio de la dinámica de la materia seca por órgano, las relaciones fuente-vertedero, las tasas fotosintética y respiratoria, la concentración de pigmentos fotosintéticamente activos, como clorofila a, b y total y los modelos de crecimiento son de gran utilidad para definir los requerimientos de regeneración de colectas y en la selección de progenitores por características de interés, para enfrentar los retos de seguridad alimentario y de cambio climático (González et al. 1995; Mohr & Schopfer, 1995).
Estudios con marcadores moleculares sugieren que las poblaciones de fríjol se clasifican en dos grupos: uno, de Meso-América, que corresponde a genotipos de semilla pequeña y, el otro, de la parte central de los Andes, que concierne a variedades de semilla grande (Gepts & Bliss, 1986; Kwak & Gepts, 2009); los dos grupos genéticos también difieren por su potencial de rendimiento. En general, las accesiones mesoamericanas se destacan por presentar mayor producción que las andinas; este comportamiento puede ser función de la rata de acumulación de biomasa durante su crecimiento (Sexton et al. 1994).
Las características para la identificación de la variabilidad genética entre los diferentes acervos genéticos de las especies silvestres y cultivadas de fríjol, casi siempre han sido fenotípicas, como el tamaño de la semilla y la forma y el tamaño de la hoja, la longitud de los entrenudos y el hábito de crecimiento, el tipo de faseolina, las aloenzimas, y el ADN del núcleo y de las mitocondrias (Gepts, 1988); sin embargo, los estudios donde se miden caracteres fisiológicos como aporte al conocimiento de la variabilidad genética son muy escasos (González et al. 1995).
La fotosíntesis es el proceso que más contribuye a la producción de biomasa; por tanto, el manejo genético y bioquímico de esta característica es especialmente importante en la producción de alimentos. Para mejorar la fotosíntesis es necesario manejar otros procesos, que permitan reducir los costos de energía y la tasa de respiración e incrementar el uso eficiente de los nutrientes, como P y N, así como realizar cambios morfológicos en la arquitectura de la planta (Foyer et al. 1995).
En varios cultivos se ha documentado que existe variación genética en la capacidad fotosintética. En fríjol, Lynch et al. (1992) observaron contrastes genéticos significativos en parámetros relacionados con la fotosíntesis entre poblaciones silvestres de diferentes regiones de origen, mientras Bressan & Pereira (2003) reportaron elevados valores de heredabilidad (> 0,50), para parámetros fisiológicos en fríjol cultivado, indicando una fuerte influencia de los factores genéticos sobre la fotosíntesis; en consecuencia, las diferencias fotosintéticas de los genotipos puede ser explotada en el fitomejoramiento.
Un mayor grosor de la hoja es usualmente asociado con alta tasa de asimilación de carbono foliar y varios estudios igualan la masa foliar específica con este grosor (White & Montes, 2005). Esta aproximación es útil porque ignora las diferencias entre el verdadero grosor, debido a la variación del contenido de agua, aire y otros factores. Muchos de los parámetros relacionados con el grosor de la hoja en fríjol presentan grandes diferencias entre líneas o genotipos, lo cual, es consistente con varios reportes que sugieren que materiales de Mesoamérica tienen mayor grosor de hoja que los andinos (White & Montes, 2005).
En este trabajo, se evaluaron parámetros de crecimiento y variables de eficiencia fotosintética en una colección de germoplasma de fríjol común (Phaseolus vulgaris L.) y se determinó el efecto de los componentes genético y ambiental, en la expresión del conjunto de las variables en estudio.
MATERIALES Y MÉTODOS
En la Corporación Colombiana de Investigación Agropecuaria (Corpoica), C.I. Tibaitatá, ubicado a 4°42' de latitud norte, 74°12' de longitud oeste y 2540m de altitud, en el Municipio de Mosquera (Cundinamarca, Colombia), se llevó a cabo un estudio de evaluación fisiológica de fríjol en dos semestres, con precipitación por ciclo de cultivo de 396 y 267mm y 13,2 y 13,5°C de temperatura diaria, en promedio, respectivamente. Los suelos de los lotes, donde se efectuó el trabajo, presentaron niveles adecuados de fertilidad, con estructura franco limosa, materia orgánica 3,7%, fósforo 30ppm, capacidad de intercambio catiónico 31,8 y meq/100g, de suelo de: Ca 19,0; Mg 2,8; K 0,19 y Na 0,63 (Flor, 1985).
Se utilizaron 29 accesiones de fríjol (Ph. vulgaris L.); 25, corresponden a cultivares regionales e introducciones de los acervos genéticos Andes y Mesoamérica, de la colección colombiana de fríjol existente en el banco de germoplasma de Corpoica en el C.I. Tibaitatá; a su vez, 14 son de crecimiento determinado arbustivo (tipo I): Antioquia 10, Antioquia 19, Antioquia 27A, Boyacá 18, Cundinamarca 148, Nariño 12, Tolima 17-2, Tolima 43, Bolivia 7, México 171, México 496, México 497, Perú 40 y Perú 154; tres de hábito indeterminado arbustivo (tipo II): Antioquia 20, Tolima 17-1 y Argentina 1 e indeterminado postrado (tipo III), ocho accesiones: Antioquia 21, Cauca 34, Huila 5, Nariño 7A , Tolima 16, Tolima 16B, Perú 5 y Perú 224. Esta colección incluyó cuatro variedades mejoradas de importancia comercial en zonas frías de Colombia: Diacol Andino, ICA Cerinza y la línea élite L-34400 de hábito tipo I e ICA Tundama de habito de crecimiento tipo II, usadas como testigos.
Las 25 accesiones regionales e introducidas, se tomaron al azar y representan el 15% de la colección de germoplasma, conformada por 165 accesiones. Todas las colectas, se sembraron en dos semestres o ambientes en la localidad C.I. Tibaitatá, bajo el diseño experimental de bloques completos al azar, con 29 tratamientos y cuatro replicaciones. Los tratamientos fueron las accesiones. La unidad experimental fue un área de 9,6m2, estuvo constituida por 4 surcos de 4m de largo, distanciados 0,6m entre ellos, con distancia de 10cm entre plantas.
La medición de las variables de peso seco y área foliar, se realizó en una planta por parcela, a partir de los 17 días después de la emergencia del cultivo hasta el estado de madurez fisiológica; las lecturas se tomaron cada 8 días, para un total de 22; se hizo la selección de plantas, con el objeto de hacer uniforme el estado biológico de las muestras.
En la tabla 1, se relacionan las 34 variables cuantitativas de origen fisiológico evaluadas en el estudio. Para tal efecto, se separaron en cada planta las hojas, los tallos, la raíz remanente después del arranque de la planta del suelo y los frutos y se secaron en un horno Mettler PE 3600, a 70°C, por 48 horas, en las que se determinó el peso seco por órgano y total de la planta; el área foliar, se midió en material fresco, empleando el medidor LI-300.
La fotosíntesis neta, se halló con el equipo portátil Licor 6200, sobre uno de los trifolios del estrato superior de la planta, en la fase vegetativa, en la etapa fenológica V4: tercera hoja trifoliada y en la fase reproductiva, en las etapas fenológicas R6: floración y R7: formación de vainas (Fernández et al. 1985). De igual manera, en extractos vegetales de plantas en etapas V4, R6 y R7, mediante espectrofotometría, se cuantificó la absorbancia a 649 y 665nm, con lo cual, a su vez, se determinó la concentración de clorofila α, β y la total.
Con base en los datos del peso seco y área foliar, se calcularon los índices de crecimiento: tasa de asimilación neta (TAN), tasa de crecimiento relativo (TCR), relación área foliar (RAF), duración área foliar (DAF), como relación de área foliar contra el tiempo entre dos momentos, área foliar específica (AFE) y relación peso foliar (RPF), por el método tradicional, propuesto por Radford (1967), con cumplimiento de los supuestos fisiológicos de variación del peso seco, en forma continua, en un período de tiempo t1a t2y dependencia lineal entre el peso seco total y el área foliar. La dinámica de la materia seca expresada en porcentaje, se calculó con base en el peso seco acumulado, a través del tiempo por cada órgano y para cada una de las 29 accesiones de fríjol.
Los datos de las variables fisiológicas, se procesaron con el sistema SAS®, versión 9,4. Se determinaron las estadísticas descriptivas, como promedios y desviaciones estándar y se realizaron los análisis de varianza para cada variable, en los dos ambientes de evaluación; con el procedimiento Univariate, se obtuvo la prueba de normalidad del supuesto del modelo estadístico (eij ≈ N I (0, σ2). Las variables de peso seco y área foliar sin normalidad, se transformaron a logaritmo natural (loge).
Los componentes de varianza, se generaron a partir de los cuadrados medios de las repeticiones y ambientes, como efectos fijos y accesiones y, el error experimental, como efectos aleatorios del análisis de varianza; este análisis, se realizó con la aplicación del procedimiento Varcomp. Los estimadores de los componentes, se calcularon de la siguiente forma:
σ2= M1; σ = (M3-M2) /r.a;
σ2ga= (M3-M2) /r;
σ2a= (M5-M3-M4+M1) /g.r
Donde: g = accesión; a = ambiente; r = repetición; y los valores de M son los cuadrados medios de cada fuente de variación.
Con estos estimadores, se halló el coeficiente de repetibilidad (Υ), propuesto por Goodman & Paterniani (1969), para estudios de clasificación de colectas, que permite determinar las variables con bajo aporte ambiental sobre la expresion fenotipica de los materiales, que a su vez, pueden ser útiles para hallar simulitud y dismilitud entre eccesiones (Ligarreto y Martinez, 2002). El coeficiente se representa como:
Υ=[σ2g/(σ2a+σ2ga) ] > 1
Los caracteres con σ>1 indican que la medida entre accesiones tiene más expresión genética que efecto de ambiente e interacciones. Los estimadores negativos de las varianzas se pueden deber a errores en el muestreo y toma de los datos causados por efecto de ambiente (Ligarreto, 2003; Chávez, 2003).
RESULTADOS Y DISCUSIÓN
Los promedios, las desviaciones estándar, las varianzas y los coeficientes de variación sobre la colección de 29 accesiones de fríjol sugieren la presencia de alta variación entre las accesiones para todas las variables, en los dos ambientes de estudio (Tabla 2). La variable que indicó menor dispersión de las accesiones fue la temperatura de la hoja, por presentar el coeficiente de variación más bajo, con valor de 3,9%, debido a que la morfología de las hojas entre las accesiones fue similar entre sí.

Un amplio rango de dispersión correspondió a las variables porcentaje de peso seco de la flor, conductancia de CO2y conductancia estomática, los valores altos de varianza y las desviaciones estándar pueden obedecer a la magnitud y al rango de las variables, como también al efecto de ambiente y a la época de muestreo (White & Montes, 2005).
Según la tabla 3 y asumiendo lo sugerido por Goodman & Paterniani (1969), que las variables con coeficientes de repetibilidad (Υ) superior a 1 aseguran su estabilidad en los diferentes ambientes de evaluación, se encontró en el estudio que las únicas variables de heredabilidad alta fueron la razón área foliar (RAF), con Υ=3,97 y la razón peso foliar (RPF), de Υ=1,38; mientras el área foliar específica (AFE), al presentar Υ=0,90, se podría considerar como de heredabilidad media; las otras 31 variables implican en su expresión diferencias entre las 29 accesiones, a través de los ambientes de evaluación, es decir, dependen en alto grado del efecto de ambiente, por presentar valores Υ cercanos a cero (Tabla 3).
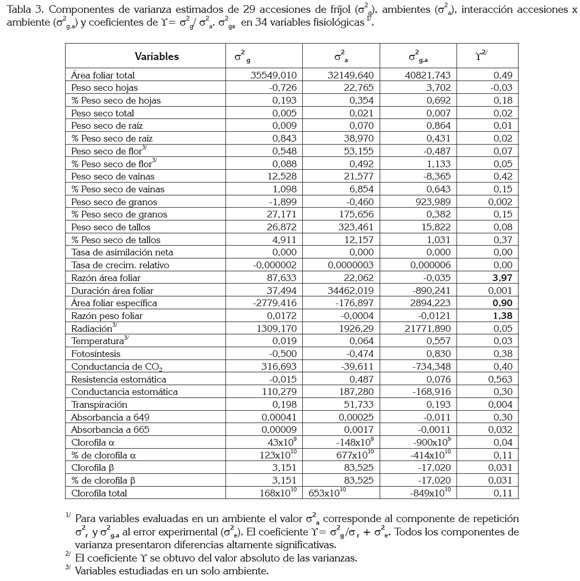
La RAF es un parámetro compuesto, parcialmente determinado por la distribución (RPF) y la morfología de la hoja (AFE) (Bayuelo et al. 2003). Se entiende la RAF como la relación del material asimilatorio por unidad de material de la planta, la RPF como la utilización del material asimilatorio para la producción de hojas y la AFE como la masa por unidad de área foliar: las tres variables están relacionadas con la tasa de fotosíntesis (White & Montes, 2005).
El alto componente genético en la expresión de estas variables implican bajo efecto del ambiente, porque son índices que relacionan dos variables de respuesta, como pesos o áreas foliares que, implícitamente, llevan este efecto, lo cual, es diferente a los índices derivados de una variable de respuesta peso o área, en relación con el tiempo (Radford, 1967).
La baja expresión genética en caracteres de peso seco y fotosíntesis en este estudio contrasta con lo obtenido por Bressan & Pereira (2003), quienes reportaron heredabilidades medias a superiores (>50%), atribuidas a la divergencia fisiológica entre los progenitores usados y a la alta expresividad de los caracteres en la población segregante. También, Ligarreto (2005), en trabajos de variabilidad en germoplasma de fríjol mediante análisis de componentes principales, determinó que la clasificación de cultivares por respuesta a 34 variables de índole fisiológica, se debe en su orden a: área foliar total, peso seco de hojas, RAF, AFE fotosíntesis y a los índices de crecimiento TAN y RPF.
De acuerdo con Poehlman & Allen (2003), los reportes de heredabilidad se aplican solamente a la población particular muestreada y al ambiente, en el cual, la población se cultivó, por ende, la interpretación de los resultados es puntual; no obstante, dado que en el estudio los materiales evaluados se cogieron al azar de la colección de 165 entradas de fríjol arbustivo, las estimaciones de heredabilidad alta para RAF, RPF y la heredabilidad media para AFE, se pueden extrapolar a toda la población muestreada, en este caso a la colección de frijol tipos I, II y III, del banco de germoplasma.
Cuando se trabaja con variabilidad producto de componentes principales lo que se tiene son varianzas acumuladas de toda la población, producto de los datos de evaluación o descripción de las plantas, por lo que se puede seleccionar las variables de mayor varianza total; en tanto, al tener varianzas de poblaciones en ambientes, se puede estimar, de manera confiable, si el carácter se hereda (Rojas, 2003).
Los resultados de este trabajo que sugieren las variables RAF, RPF y AFE con aporte genético importante en la clasificación de accesiones de germoplasma de una colección colombiana de fríjol por parámetros fisiológicos, coinciden con varios reportes, que las consideran como variables de gran utilidad en los análisis de crecimiento, al estar estrechamente relacionadas con la tasa potencial de crecimiento de las plantas (Lambers & Poorter, 1992), además de estar asociadas con una mayor tasa de asimilación de carbono (White & Montes, 2005).
En los dos acervos genéticos de fríjol andino y mesoamericano, la RAF presentó tendencia creciente a través del tiempo, con picos altos entre prefloración (R6) y floración (R7), cuando la plantas alcanzaron su mayor crecimiento foliar y el desarrollo de sus estructuras reproductivas. Para el efecto, en la figura 1:A, B, se presenta el comportamiento promedio de la RAF por pool genético, con las accesiones L-34400 y Cauca 34, representativas de cada acervo, como lo indica Ligarreto (2005). En cambio, para el uso del material asimilatorio en la producción de hojas (RPF), el promedio del ciclo de vida de la planta fue mayor en el acervo mesoamericano que en el andino, comportamiento que pudo obedecer a la mayor área foliar de las accesiones mesoamericanas, con predominio de hábito de crecimiento indeterminado postrado (tipo III), lo que a su vez, coincide con lo reportado por Maldonado & Corchuelo (1993), quienes detectaron mayor disminución de RPF durante el ciclo de vida, en variedades de fríjol de hábito determinado (I), con respecto al hábito de crecimiento indeterminado (II y III).
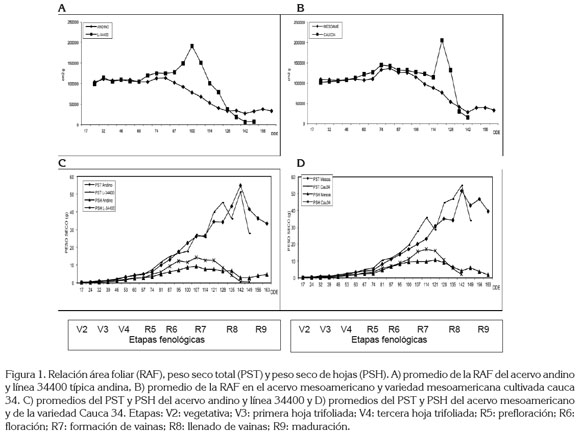
Los puntos máximos de incremento de la RAF en las dos accesiones L-34400 y Cauca 34, obedecen a puntos de inflexión en el peso seco total de la planta, atribuidos al comienzo de la traslocación de fotoasimilados y a la remoción de elementos constituyentes del tejido foliar de las plantas hacia las vainas y los granos, con el fin de contribuir a su llenado (Figura 1), es decir, que el suministro de asimilados afecta poco el grosor de la hoja. El fríjol es una planta C3 con rápida traslocación de asimilados, debido a las cortas distancias entre la fuente y la demanda, en especial, cuando los cultivares son de hábito arbustivo (González et al. 1995; Ligarreto, 2005).
El grosor de las hojas (AFE) promedio de ciclo de vida del cultivo, para el caso específico de las accesiones L-34400 y Cauca 34 representativas de los acervos andino y mesoamericano, respectivamente, y con hábitos de crecimiento I y III, en su orden fue de 374,34 y de 179,78cm2g-1; la mayor AFE en la accesión del hábito de crecimiento determinado (I) fue ocasionada por la reducción en el peso de las hojas, como también a la translocación de asimilados hacia las estructuras reproductivas (figura 1 A y C). Caso contrario es el de la accesión Cauca 34 con menor AFE, posiblemente, al incremento del peso de las hojas al continuar la producción de las mismas, por su condición de hábito indeterminado, ocurriendo la fase vegetativa al mismo tiempo que la reproductiva y la translocación de asimilados hacia las vainas y nuevas hojas, ocurre de manera simultánea (Ligarreto, 2005).
Según Joshi (1997), los reportes acerca de la heredabilidad de los factores que determinan la biomasa y la fotosíntesis en fríjol común son escasos, por ser procesos complejos, controlados por un gran número de genes y estar fuertemente afectados por el ambiente; efecto que también se encontró en este estudio, al presentar muchas de estas variables coeficientes de repetibilidad menores a la unidad (Υ<1).
Rosales et al. (2004), en estudios de cultivares de fríjol, detectaron diferencias en la acumulación y en la distribución de biomasa por hábitos de crecimiento. En este mismo sentido, la fotosíntesis, aún cuando tiene baja heredabilidad, por ser el proceso que más contribuye a la formación de biomasa, se ha considerado útil en algunos estudios de variabilidad genética y en la selección de progenitores y líneas de mejoramiento.
A partir del valor agregado de la colección por estos índices de crecimiento, se pueden determinar agrupamientos de accesiones para realizar acciones de enriquecimiento de las bases genéticas del germoplasma y así poder ofertar progenitores con alelos de interés para programas de fitomejoramiento. Cuando se desee trabajar poblaciones de fríjol para selección por estas variables, se pueden usar los métodos de mejoramiento masal o selección recurrente, que son adecuados para el manejo de variables cuantitativas al incrementar, de manera gradual, la frecuencia de los alelos favorables, sin reducir la variabilidad genética de la población inicial (Poehlman & Allen, 2003).
De acuerdo con Bayuelo et al. (2002), en fríjoles cultivados (Ph. vulgaris y Ph. acutifolius), factores adversos del cultivo causan estrés en las plantas reduciendo la RAF, por consiguiente, es pertinente realizar selección de progenitores en los bancos de germoplasma, dado su comportamiento por este índice. En el caso de cultivares susceptibles al factor adverso salinidad de los suelos, se puede presentar menor RAF causada por un decrecimiento en la AFE, al haber un incremento de biomasa por unidad de área foliar, como efecto de ajuste osmótico de la hoja, debido a una elevada concentración de sal o un decrecimiento en la proporción de materia seca distribuida en el tejido foliar (RPF).
Según Bayuelo et al. (2003), en cultivares de fríjol la RPF mantiene valores relativamente estables, con incrementos de la salinidad entre 0,40 y 80mM de NaCl; mientras la AFE decrece significativamente con el estrés a la salinidad, igual comportamiento presenta la RAF, indicando a su vez, la alta sensibilidad hacia el desarrollo del área foliar en fríjol. El decrecimiento de RAF con la salinidad fue, en primer lugar, debido al decrecimiento en la AFE (incremento en biomasa por unidad de área), antes que al decrecimiento en la proporción de distribución de materia seca en el tejido foliar (RPF).
En fríjol cultivado y silvestre resulta importante considerar el comportamiento de los índices de crecimiento AFE, RPF y AFE y su asociación con el peso seco de las estructuras y el total de la planta y con el área foliar, por ser variables derivadas de estos caracteres. Su valoración aporta al conocimiento de la distribución de recursos desde la zona foliar hacia las otras estructuras reproductivas y de anclaje, como la raíz. Además, permite discriminar a los cultivares por parámetros de índole fisiológico.
La afirmación que las variables fisiológicas son procesos complejos, controlados por muchos genes y con fuerte afectación por el ambiente, fue confirmada para un gran número de variables, como peso seco de estructuras, área foliar y fotosíntesis en fríjol, con excepción en los índices de crecimiento RAF, RPF y AFE, que representaron estabilidad al ambiente y expresaron aporte genético útil para estudios de medición de caracteres en poblaciones, tanto para analizar diversidad genética en programas de recursos fitogenéticos como para procesos de selección de progenitores, en el premejoramiento genético y líneas de mejora genética de la especie.
Puede resultar útil la hibridación de los dos acervos genéticos presentes en la colección colombiana de fríjol para generar incrementos en rendimiento, con el aporte en la eficiencia fisiológica por parte de los cultivares mesoamericanos y la adaptación a las zonas productoras, por el acervo andino. Hay que considerar que se puede tener avance genético al realizar cruces dentro de cada acervo dada la gran variabilidad genética existente en cada pool genético.
Agradecimientos: El autor agradece a la División de Investigación de la Universidad Nacional de Colombia, Bogotá y a Colciencias, por el soporte financiero. Al profesor Stanislav Magnitskiy, de la Facultad de Agronomía, Universidad Nacional de Colombia, sede Bogotá, por la revisión del manuscrito. Conflicto de intereses: El manuscrito fue preparado y revisado por el autor, quien declara que no existe ningún conflicto de intereses que ponga en riesgo la validez de los resultados presentados.
BIBLIOGRAFÍA
1. BAYUELO, J.S.; DEBOUCK, D.G.; LYNCH, J.P. 2002. Salinity tolerance of Phaseolus vulgaris species during early vegetative growth. Crop Sci. 42:2184- 2192.
2. BAYUELO, J.S.; DEBOUCK, D.G.; LYNCH, J.P. 2003. Growth, gas exchange, water relations, and ion composition of Phaseolus species grown under saline conditions. Field Crops Res. 80:207-222.
3. BRESSAN, R.; PEREIRA M.G. 2003. Inheritance analysis of photosynthetic characteristics in bean (Phaseolus vulgaris L.). Physiol. Mol. Biol. Plants. 9(2):249-253.
4. BORÉM, A. 1998. Melhoramento de plantas. 2ª ed. Vicosa, Editora UFV. 453p.
5. BROUGHTON, W.J.; HERNANDEZ, G.; BLAIR, M. 2003. Beans (Phaseolus spp.) - model food legumes. Plant and Soil. 252:55-128.
6. CHÁVEZ, J.L. 2003. Análisis estadístico de datos de caracterización morfológica. p.72-76. En: Franco, T.; Hidalgo, R. (eds). Análisis estadístico de datos de caracterización morfológica de recursos filogenéticos. Bol. Técn. IPGRI n. 8. Roma, Italia, v.8, 89p.
7. FAO. 2011. FAOSTAT. Disponible desde Internet en: http://faostat.fao.org/site/567/default.aspx#ancor (con acceso 05/12/2011).
8. FERNÁNDEZ, F.; GEPTS, P.; LÓPEZ, M. 1985. Etapas de desarrollo en la planta de fríjol. p.61-78. En: López, M., Fernández, F.; Van Schoonhoven, A. (eds.). Fríjol investigación y producción. CIAT, Cali, Colombia.417p.
9. FLOR, C. 1985. Revisión de algunos criterios sobre la recomendación de fertilizantes en frijol. En: López, M., Fernández, F.; Van Schoonhoven, A. (eds). Fríjol investigación y producción. CIAT, Cali, Colombia. p.287-312.
10. FOYER, C.; CHAUMEONT, M.; MURCHIE, E.; GALTIER, N.; FERRARIO, S. 1995. End-product modulation of carbon partitioning with a view to improved biomass production. In: Madore, M.A.; Lucas, W.J. (eds). Carbon parttioning and source-sink interactions in plants. Am. Soc. Plants Physiol. p.45-55.
11. GEPTS, P.; BLISS, F.A. 1986. Phaseolin variability among wild and cultivated common beans (Phaseolus vulgaris) from Colombia. Econ. Bot. 40(4):469-478.
12. GEPTS, P. 1988. Genetic resources of Phaseolus beans. Kluwer Acad. Publish. p.215-241.
13. GONZÁLEZ, A.; LYNCH, J.; TOHME, J.M.; BEEBE, S.E.; MACCHIAVELLI, R.E. 1995. Characters related to leaf photosynthesis in wild populations and landraces of common bean. Crop Science 35:1468-1476.
14. GOODMAN, M.M.; PATERNIANI, E. 1969. The races of maize III choices appropriate characters for racial classification. Econ. Bot. 23:265-273.
15. JARVIS, A.; RAMIREZ, J.; BONILLA-FINDJI, O.; ZAPATA, A. 2011. Impacts of climate change on crop production in Latin America p. 44-56. In: YADAV, S.S.; REDDEN, R.; HATFIELD, J.L.; LOTZE- CAMPEN, H.; HALL, A. (eds.) Crop Adaptation to Climate Change. Wiley-Blackewll, Oxford, UK. 596p.
16. JOSHI, A.K. 1997. Genetic factors affecting photosynthesis. In: Handbook of photosynthesis, Ed. Pessarakli, M.; Dekker, M. (eds). Handbook of Photosynthesis. p.751-767.
17. KWAK, M.; GEPTS, P. 2009. Structure of genetic diversity in the two major gene pools of common bean Phaseolus vulgaris L., Fabaceae). Theor. Appl. Genet. 118:979-992.
18. LAMBERS, H.; POORTER, H. 1992. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Adv. Ecol. Res. 23:187-261.
19. LIGARRETO, G.A. 2003. Conceptos y mediciones útiles para la caracterización de germoplasma. p.77-79. En: Franco, T.; Hidalgo, R. (eds). Análisis estadístico de datos de caracterización morfológica de recursos filogenéticos. Bol. Técn. IPGRI no.8. Roma, Italia, v.8, 89 p.
20. LIGARRETO, G.A. 2005. Uso de índices de crecimiento y caracteres relacionados con la fotosíntesis para el análisis de la variabilidad genética de fríjol común (Phaseolus vulgaris L.). Fitotec. Col. 5(1):23-35.
21. LIGARRETO, G.A.; MARTÍNEZ, O. 2002. Variabilidad genética en fríjol común (Phaseolus vulgaris L.), análisis de variables morfológicas y agronómicas cuantitativas. Agr. Col. 19(1-2):69-80.
22. LYNCH, J.; GONZÁLEZ, A.; TOHME, J.M.; GARCIA, J.A. 1992. Variation in characters related to leaf photosynthesis in wild bean populations. Crop Sci. 32:633-640.
23. MALDONADO, G.; CORCHUELO, G. 1993. Dinámica del crecimiento de dos variedades de frijol (Phaseolus vulgaris L.). Agr. Col. 10(2):114-121.
24. MOHR, H.; SCHOPFER, P. 1995. Plant Physiology. Springer-Verlag Berlin Heidelberg New York. 629p.
25 POEHLMAN, J.M.; ALLEN, D. 2003. Mejoramiento genético de las cosechas. Ed. Limusa, 2aed. México, D.F. 511p.
26. RADFORD, P.J. 1967. Growth analysis formulae-their use and abuse. Crop Sci. 7(3):171-174.
27. ROJAS, W. 2003. Caracterización morfológica de germoplasma. Estudio de caso 1. Análisis de la variabilidad genética en quinua, p 27-39. En: Franco, T.; Hidalgo, R. (eds). Análisis estadístico de datos de caracterización morfológica de recursos filogenéticos. Bol. Técn. IPGRI n°.8. Roma, Italia, 89 p.
28. ROSALES, R.; KOHASHI, J.; ACOSTA, J.A.; TREJO, C.; ORTIZ, J.; KELLY, J.D. 2004. Biomass distribution, maturity acceleration and yield in drought-stressed common bean cultivars. Field Crops Res. 85:2003- 2011.
29. SEXTON, P.J.; WHITE, J. W.; BOOTE, K.J. 1994. Yield- determining processes in relation to cultivar seed size of common bean. Crop Sci. 34:84-91.
30. SHAPIRO, S.S.; WILK, M.B. 1965. An analysis of variance test for normality (complete samples). Biometrika. 52:591-611.
31. WHITE, J.W.; MONTES, C. 2005. Variación in parameters related to leaf thickness in common bean (Phaseolus vulgaris L.). Field Crops Res. 91:7-21.
Recibido: Julio 30 de 2012 Aceptado: Febrero 28 de 2013

Revista U.D.C.A Actualidad & Divulgación Científica por Universidad de Ciencias Aplicadas y Ambientales se distribuye bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.